Clear Sky Science · zh
对COMATOSE转运蛋白的预测建模揭示了保守的酰基-CoA配位口袋
植物如何将储存的脂肪转化为燃料
当种子苏醒并开始生长时,必须迅速将储存的油脂转化为可用的能量。该过程中的关键角色是名为COMATOSE的膜上“闸门”,它有助于将脂类分子输送到可被分解的小细胞隔室中。本研究利用强大的计算工具预测COMATOSE的三维结构和运作周期,揭示它如何抓取并移动货物——以及类似转运蛋白在人体健康与疾病中的工作方式。
脂类燃料的微观通道
在大多数植物和动物细胞内,小囊泡状的过氧化物酶体负责处理某些脂类分子并将其转化为有用的构件。为此,它们依赖于一类称为ABCD转运蛋白的家族。在模式植物拟南芥中,只有一个过氧化物酶体ABCD转运蛋白,称为COMATOSE(CTS),它对种子萌发至关重要,因为它将与辅因子CoA结合的脂肪酸运入过氧化物酶体。若无CTS,种子无法正确利用其能量储备,除非外加糖分否则不能生长。尽管已有多年的遗传学和生化研究,CTS的精细结构及其脂类货物经过的确切路径仍不清楚。
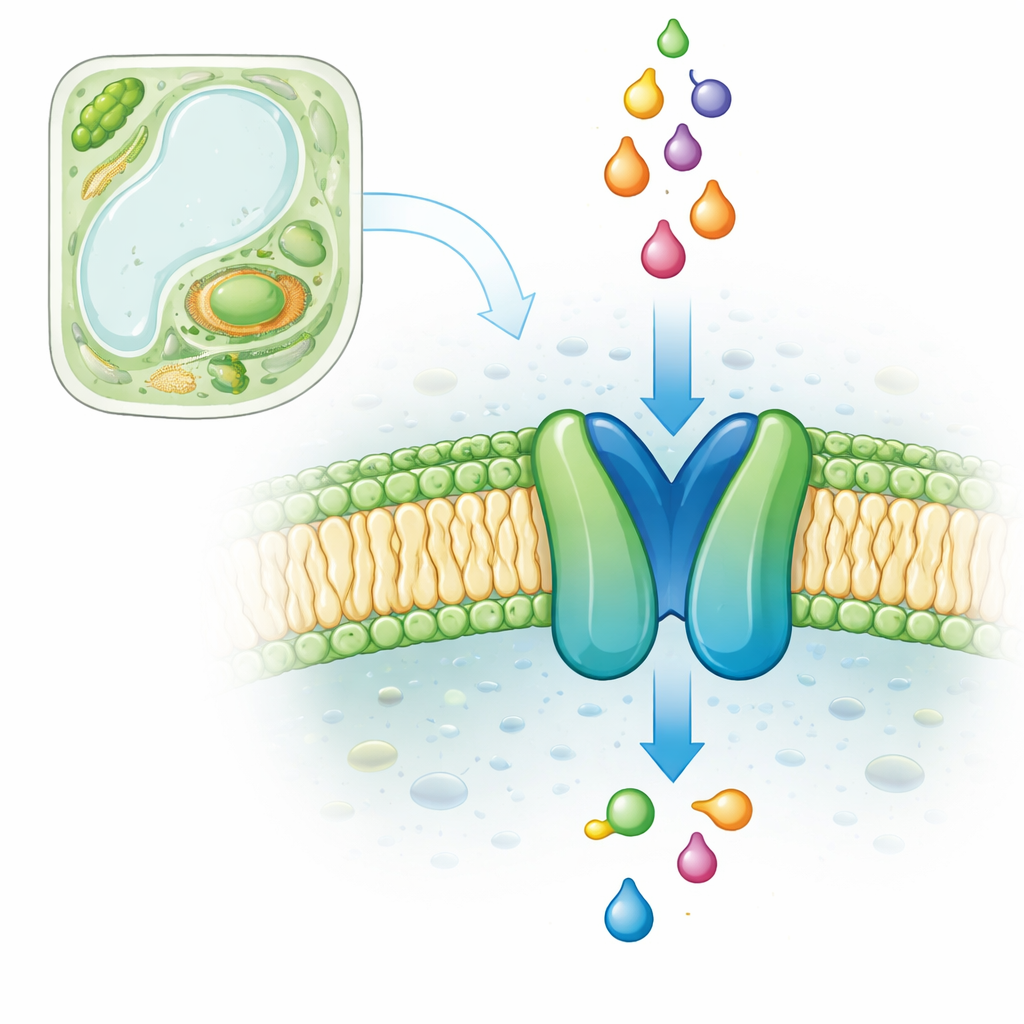
用人工智能预测蛋白形状
像X射线晶体学和冷冻电镜这样的实验方法在面对大型、柔性的膜蛋白(如CTS)时存在困难。因此,作者们转向最先进的结构预测软件AlphaFold2和AlphaFold3,这些工具利用深度学习从氨基酸序列推断三维形状。他们生成了CTS在若干“构象”下的高置信度模型:空置形式、与ATP结合的形式,以及ATP被消耗后与ADP结合的形式。这些模型显示CTS具有相关ABC转运蛋白的典型结构:两组跨膜螺旋束形成中央腔体,并与两个能量结合单元耦合,这些单元在运输周期中会合拢和分开。
寻找隐藏的货物口袋
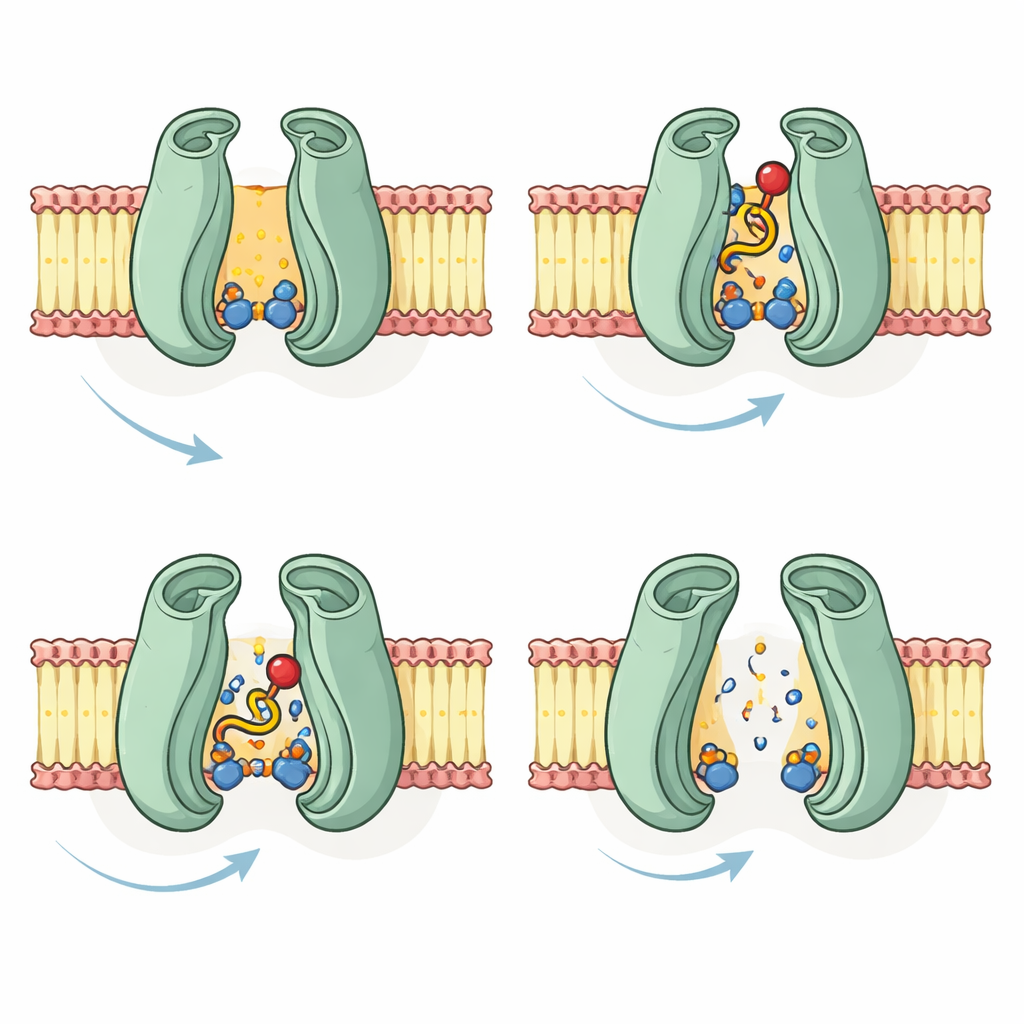
有了这些预测结构,团队模拟了长链酰基与CoA连接的脂肪酸,以及仅CoA部分,如何塞入CTS。在空(apo)模型中,这两种分子都落入位于膜区深处的同一口袋,周围被一簇带电和极性氨基酸包围。有若干残基与CoA头部形成紧密接触,提示它们帮助固定带负电的磷酸和糖。当作者在结合ATP或ADP的模型上重复对接时,脂类分子不再适配中央腔体,而是粘附在外表面,且相互作用较弱。这支持了这样一种观点:内在口袋在静息态可接近,但一旦能量被使用并准备释放货物时,该口袋会塌缩或发生位移。
跨物种的保守特征
为了检验其预测口袋是否具有广泛重要性,研究人员考察了植物及其它生物中许多相似转运蛋白各氨基酸位置的保守程度。与CoA接触的口袋内的残基被发现高度保守,暗示它们在底物识别中具有共同作用。研究团队随后将CTS模型与一份高分辨率的人类ABCD1转运蛋白与相似酰基-CoA结合的结构对齐。令人震惊的是,CoA头部的位置和环境在两种蛋白中几乎相同。这一高度吻合支持了一个从植物到人类都被保留的配位口袋的存在,并有助于解释为何这些转运蛋白中相似的突变会对代谢产生强烈影响。
重新思考先前提出的催化三联体
先前的研究曾提出CTS中一组三个保守残基可能协同作为化学“刀片”切断脂肪酸与CoA之间的键。将这些残基映射到新模型上显示,它们距已结合的CoA约有28埃——距离过远,无法直接完成该反应。相反,研究指出另一处残基——靠近结合口袋的一位丝氨酸——更有可能参与断键过程,可能由附近能激活其反应性基团的伙伴辅助。该分析还为若干已知CTS突变体的行为提供了结构学解释,这些突变要么阻断种子萌发,要么选择性地损害脂肪酸分解而保留其他活性。
这对种子及更广泛领域的意义
总体而言,该研究提出了COMATOSE的逐步运输循环:空置蛋白面向细胞内侧,接受一个或多个酰基-CoA分子进入一个保守口袋,随后在ATP结合时将能量结合单元合拢以翻转货物穿过膜并在过氧化物酶体内释放,可能在此之前切下CoA基团。尽管这些结论基于计算而非直接成像,但它们与现有的实验数据以及人类ABCD转运蛋白的结构相吻合。对于非专业读者,关键结论是:先进的蛋白结构预测现在可以在我们能通过实验捕捉到这些分子机器之前揭示它们的工作方式,从而指导未来的实验验证并加深我们对植物与人类能量利用的理解。
引用: Bifsa, F., Simmons, K., Muench, S.P. et al. Predictive modelling of the COMATOSE transporter reveals a conserved ligand binding pocket for acyl-CoAs. Sci Rep 16, 10423 (2026). https://doi.org/10.1038/s41598-026-39225-9
关键词: 过氧化物酶体转运, 脂肪酸代谢, ABC转运蛋白, 蛋白质结构预测, 植物种子萌发