Clear Sky Science · en
Predictive modelling of the COMATOSE transporter reveals a conserved ligand binding pocket for acyl-CoAs
How plants turn stored fats into fuel
When a seed wakes up and starts to grow, it must quickly turn its stored oils into usable energy. A key player in this process is a membrane "gate" called the COMATOSE transporter, which helps move fatty molecules into tiny cell compartments where they can be broken down. This study uses powerful computer tools to predict the 3D shape and working cycle of COMATOSE, revealing how it may grip and move its cargo—and how similar transporters work in human health and disease.
A microscopic doorway for fatty fuels
Inside most plant and animal cells, small sacs called peroxisomes help dispose of certain fatty molecules and convert them into useful building blocks. To do this, they rely on a family of transporter proteins known as ABCD transporters. In the model plant Arabidopsis, there is a single peroxisomal ABCD transporter, called COMATOSE (CTS), that is vital for seed germination because it imports fatty acids linked to a helper molecule called CoA. Without CTS, seeds cannot properly tap into their energy reserves and fail to grow unless extra sugar is provided. Despite years of genetic and biochemical work, the detailed structure of CTS and the exact path taken by its fatty cargo have remained unclear.
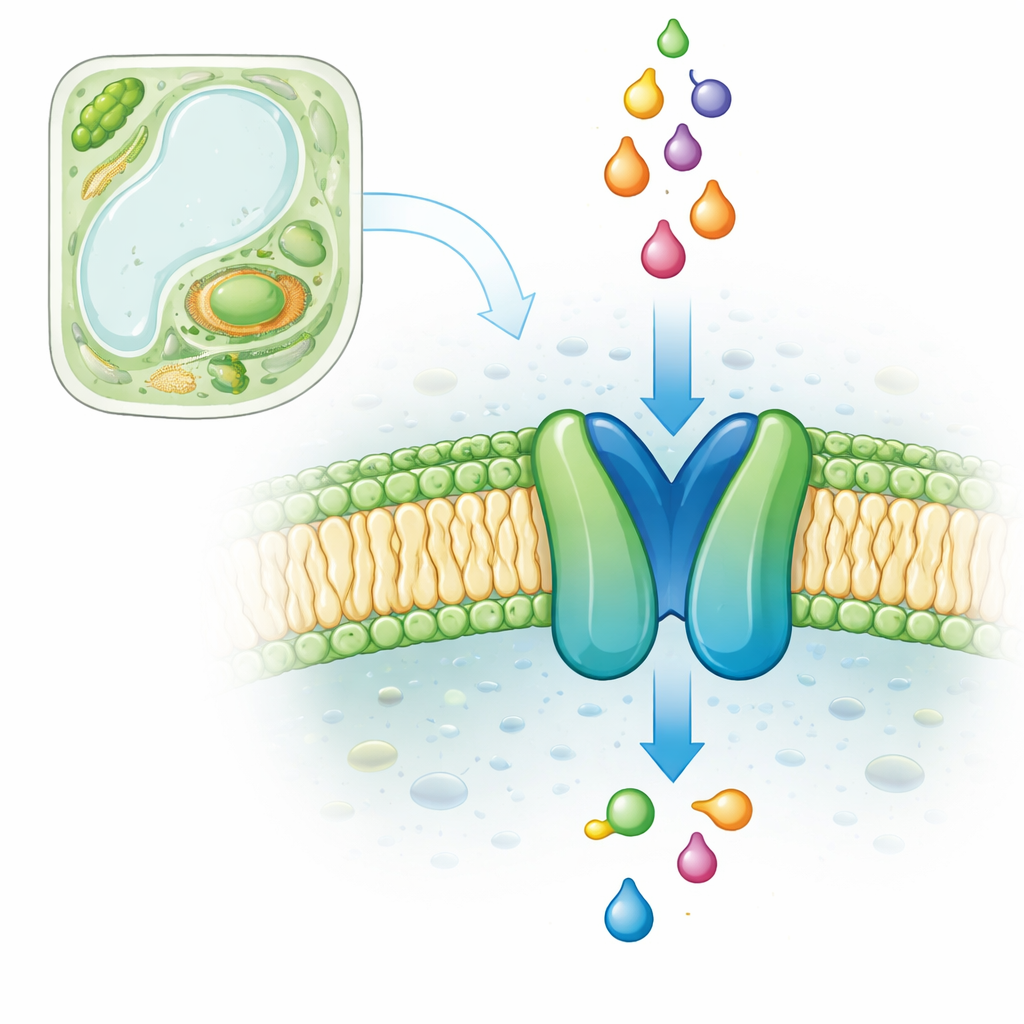
Using AI to predict protein shapes
Experimental methods such as X-ray crystallography and cryo-electron microscopy struggle with large, flexible membrane proteins like CTS. The authors therefore turned to state-of-the-art structure prediction software, AlphaFold2 and AlphaFold3, which use deep learning to infer 3D shapes from amino-acid sequences. They generated high-confidence models of CTS in several "poses": an empty form, a form bound to ATP (the fuel that powers transport), and a form bound to ADP after ATP has been used. These models showed CTS has the characteristic architecture of related ABC transporters: two bundles of membrane-spanning helices that form a central cavity, coupled to two energy-binding units that clamp together and pull apart during the transport cycle.
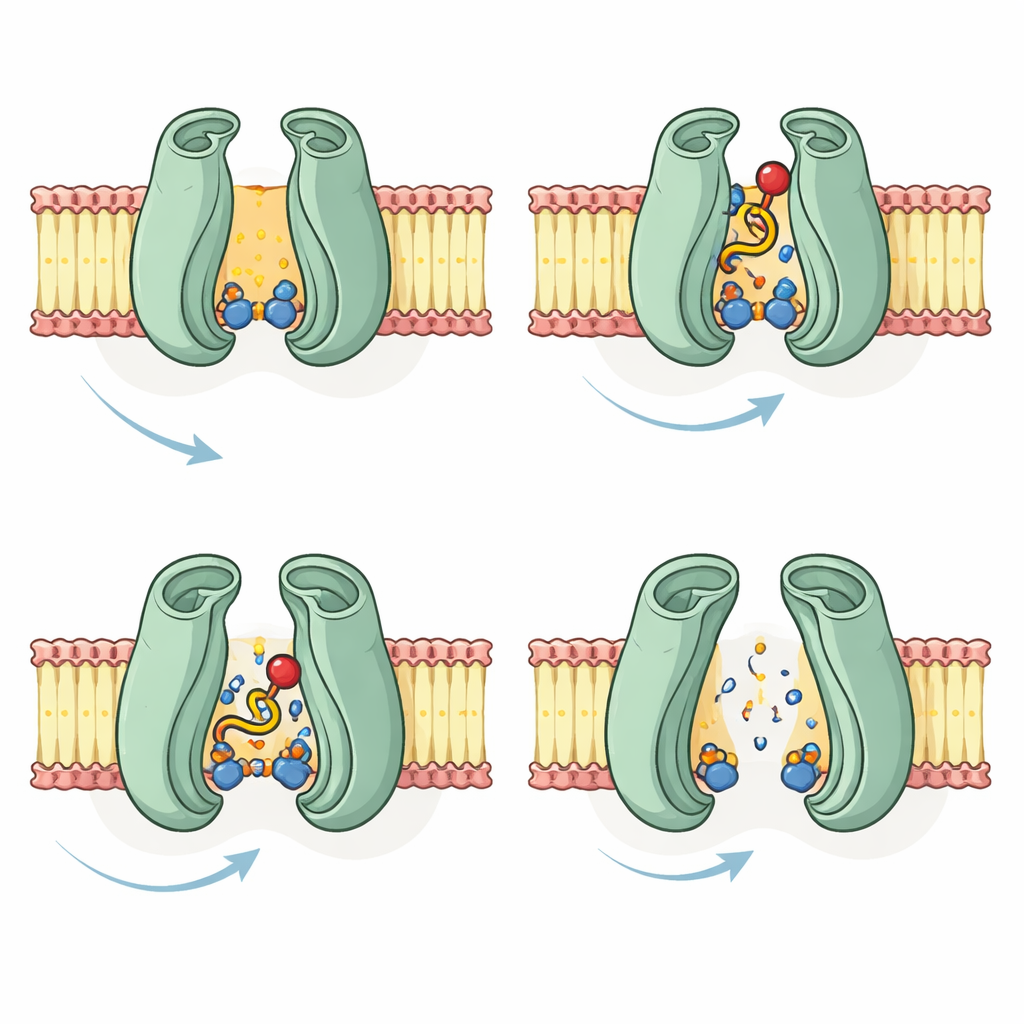
Finding the hidden pocket for cargo
With these predicted structures in hand, the team simulated how a very long-chain fatty acid attached to CoA, as well as the CoA portion alone, might fit into CTS. In the empty (apo) model, both versions settled into the same pocket deep within the membrane region, surrounded by a cluster of charged and polar amino acids. Several residues formed close contacts with the CoA headgroup, suggesting they help anchor the negatively charged phosphates and sugar. When the authors repeated the docking with models in which ATP or ADP was bound, the fatty molecules no longer fit into the central cavity and instead stuck to outer surfaces, with weaker interactions. This supported the idea that the internal pocket is accessible in the resting state, but collapses or shifts once energy has been used and the cargo is ready to be released.
Conserved features across species
To test whether their predicted pocket was broadly important, the researchers examined how strongly each amino-acid position is conserved across many similar transporters in plants and other organisms. Residues lining the CoA-contacting pocket turned out to be highly conserved, implying a shared role in substrate recognition. The team then aligned the CTS model with a high-resolution structure of the human ABCD1 transporter bound to a similar fatty acyl-CoA. Strikingly, the location and environment of the CoA headgroup were almost the same in both proteins. This close match supports the existence of a conserved binding pocket that has been maintained from plants to humans, and helps explain why similar mutations in these transporters can have strong effects on metabolism.
Rethinking a proposed catalytic trio
Previous work had suggested that a group of three conserved residues in CTS might act together as a chemical "blade" to cut the bond between fatty acid and CoA. Mapping these residues onto the new models showed that they lie about 28 angstroms away from the bound CoA—too far to carry out this reaction directly. Instead, the study points to another residue, a serine near the binding pocket, as a more plausible participant in bond breaking, possibly helped by nearby partners that can activate its reactive group. The analysis also offers structural explanations for the behavior of several known CTS mutants that either block seed germination or selectively impair fatty-acid breakdown while leaving other activities intact.
What this means for seeds and beyond
Overall, the work proposes a step-by-step transport cycle for COMATOSE: the empty protein faces the cell interior, accepts one or more fatty acyl-CoA molecules into a conserved pocket, then clamps its energy-binding units together upon ATP binding to flip the cargo across the membrane and release it inside the peroxisome, possibly after cutting off the CoA group. While these conclusions are based on computation rather than direct imaging, they fit well with existing experimental data and with structures of human ABCD transporters. For non-specialists, the key takeaway is that advanced protein-structure prediction can now reveal how crucial molecular gates in cells may work long before we can capture them experimentally, guiding future lab tests and deepening our understanding of energy use in both plants and people.
Citation: Bifsa, F., Simmons, K., Muench, S.P. et al. Predictive modelling of the COMATOSE transporter reveals a conserved ligand binding pocket for acyl-CoAs. Sci Rep 16, 10423 (2026). https://doi.org/10.1038/s41598-026-39225-9
Keywords: peroxisomal transport, fatty acid metabolism, ABC transporters, protein structure prediction, plant seed germination