Clear Sky Science · zh
线性RAG扫描介导Igκ可变区库的编辑
我们的免疫系统如何编辑自身工具
每天,我们的免疫系统都必须分辨己与非己。B细胞——制造抗体的白细胞——有时会构建出意外识别自身组织的受体。为维持健康,这些细胞需要一种内在机制来悄然修复或丢弃此类高风险受体。本研究揭示了小鼠B细胞如何在抗体基因的一部分上进行精确的“编辑”过程,重新塑造其防御工具的同时避免自体攻击。
从基因片段构建抗体部件
抗体由两大部分组成:重链和轻链,每一部分都由许多小的DNA片段组装而成。在发育中的B细胞中,一个切接机器RAG会剪切并连接这些片段,以产生多样的受体。本研究聚焦于κ轻链,其由一百多个可变片段和少数连接片段组成,分布在超过三百万个DNA碱基位点上。首次组装遵循一种构型:通过DNA环状结构将远端的可变片段带近中央的连接位点。在这一“初级”阶段,正向和反向取向的片段都可以被匹配,且一些特别强的靶序列使切接效率更高。
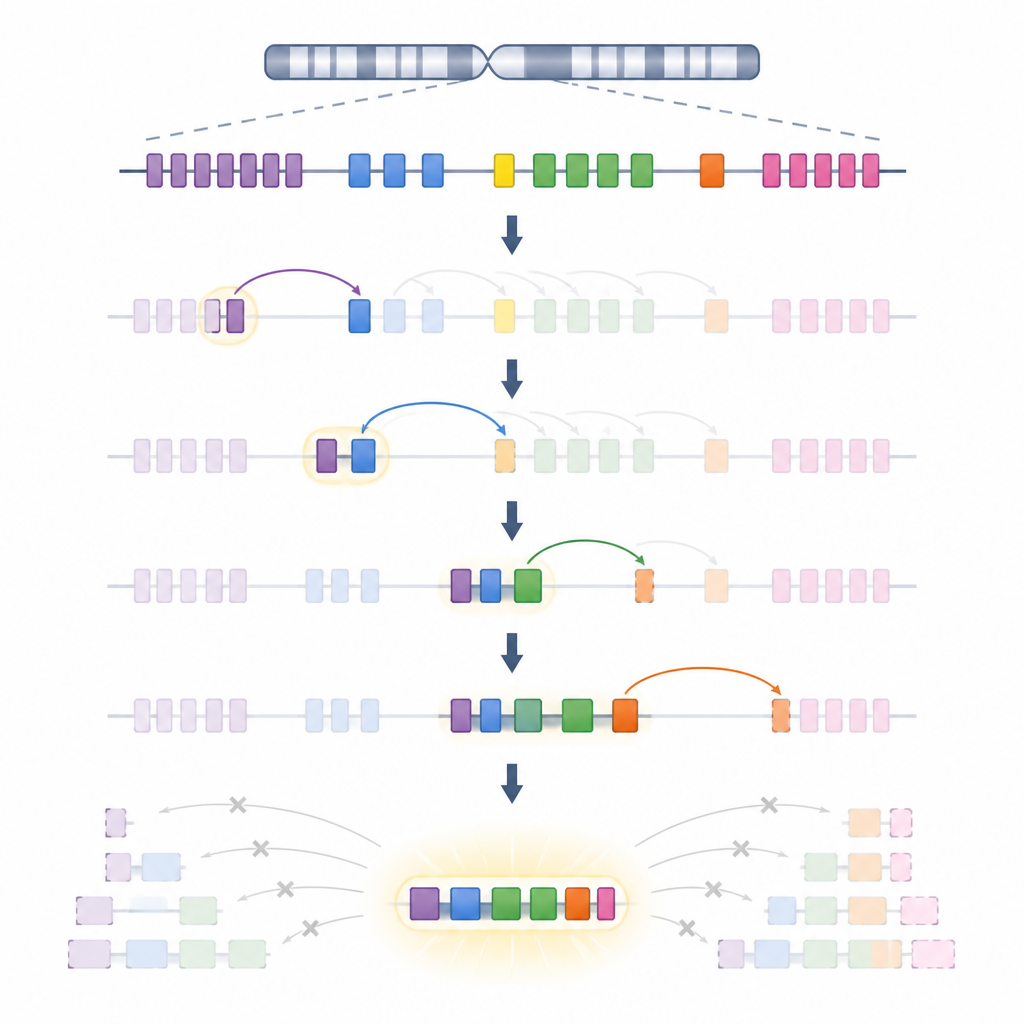
从环状结构切换到扫描模式
如果首轮轻链有缺陷或识别自身,B细胞可以在“次级”编辑中重试。作者表明,这种转换由初次连接移除或移动一个称为Cer/Sis的特殊DNA平台触发。一旦该平台丢失,RAG便不再依赖两个DNA环,而更像是沿着文本行滑动的读取器。从初次连接产生的许多新位点出发,RAG沿染色体朝一个方向扫描,按顺序测试邻近的可变片段。在细胞群体中,多个这样的扫描中心分布在轻链区域,因此尽管每个中心只搜索有限的区段,但它们共同仍能几乎采样到整个片段库。
为何邻近片段主导编辑
借助高通量的DNA连接映射与经工程改造的鼠细胞系,研究人员发现次级编辑主要使用位于每个新扫描中心上游的可变片段。两大因素使过程偏向这些邻近片段。其一,当某个可变片段被积极转录为RNA时,该局部转录活性似乎会减慢扫描机器的移动,使RAG与该片段接触更多。其二,某些可变片段携带异常强的靶序列,对RAG有高度吸引力。局部RNA产生与强靶序列共同作用,使邻近片段被优先且频繁使用,快速“饱和”可能的连接,从而限制扫描器通常能行进的距离。
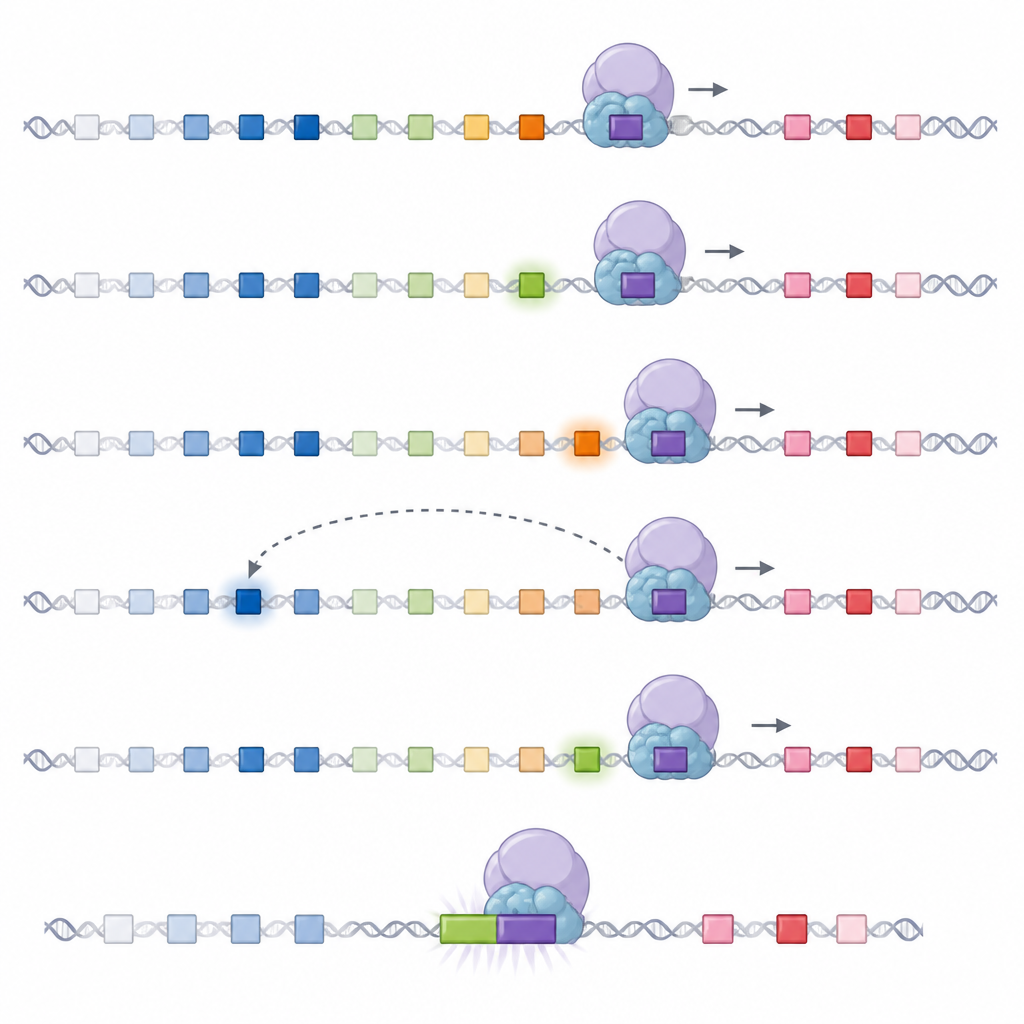
允许有限的回溯与翻转
研究还探讨了当某些可变片段处于“错误”取向时会发生什么。在重链基因中,此类反向片段在扫描过程中几乎不会被使用。然而在此处,作者表明对于轻链,强靶序列可以允许反向片段在次级阶段发生连接,要么通过真实的翻转(inversion),要么通过类似翻转的缺失过程。通过在细胞模型中精心重设计这些信号,他们证明只有强信号支持这种不寻常的连接,并且这仍符合同一基于扫描的框架,有时伴随一小段局部运动以对齐片段。
对免疫平衡的意义
总体而言,这些发现表明B细胞在初次基于环状结构的组装步骤之后,通过受控的单向扫描过程来编辑其κ轻链。该切换使细胞能够利用一组集中且邻近的基因片段修复或替换有风险的受体,同时保证在大量细胞中仍可触及轻链区的整体多样性。对非专业读者而言,关键信息是:抗体基因并非一次性固定——它们内置了一套“查找并替换”系统,既能细致调整对外来靶标的识别,又有助于防止对自身造成有害反应。
引用: Li, X., Hu, H., Zhang, Y. et al. Linear RAG scanning mediates editing of Igκ variable region repertoires. Nature 653, 870–878 (2026). https://doi.org/10.1038/s41586-026-10362-5
关键词: B细胞发育, 抗体基因重排, 受体编辑, 免疫耐受, RAG扫描