Clear Sky Science · zh
勒普廷缺乏的肥胖小鼠绝食期间代谢反应能力的全面丧失与酶水平升高
为什么饥饿时肌肉很重要
当我们错过一顿饭时,身体会迅速重排能量的使用与产生方式,以确保重要器官,尤其是大脑,继续运作。但在肥胖状态下,这种紧急重排常常出现故障。本研究提出了一个简单却重要的问题:当食物突然消失时,肥胖动物的肌肉和肝脏与瘦动物有何不同?这些差异说明了它们的健康状况何种问题?通过同时追踪数千种分子,研究人员揭示了肥胖中一种隐秘的代谢“敏捷性”丧失,这可能有助于解释胰岛素抵抗和减重困难等问题。
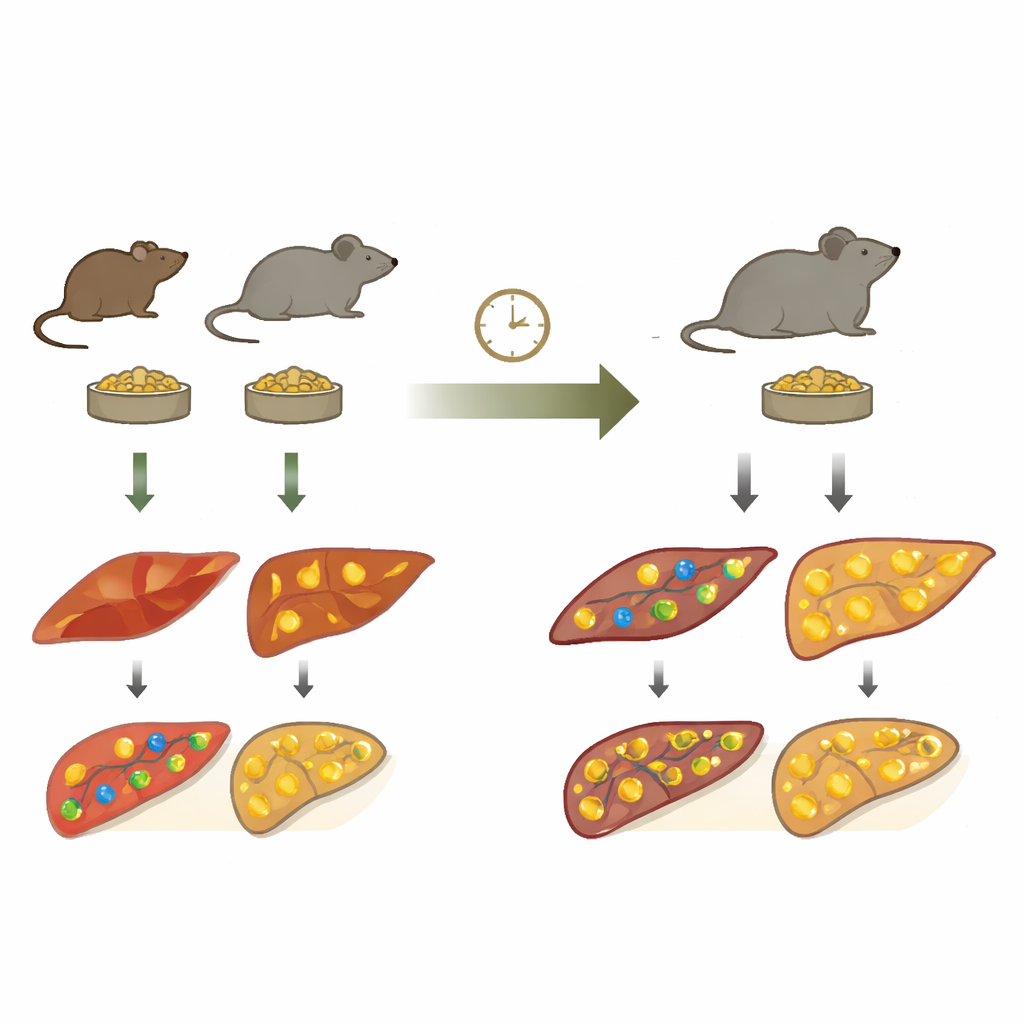
两类小鼠的故事
科学家将正常小鼠与勒普廷缺乏的“ob/ob”小鼠进行比较,后者是一个经典的严重肥胖基因模型。两组均被禁食24小时,并在八个时间点采集骨骼肌和肝脏样本。研究团队没有只观察某一类分子,而是测量了小分子代谢物、基因表达(RNA)、蛋白质水平以及蛋白质磷酸化——细胞开关酶活性的常见方式。随后他们将这些层次拼接成“跨组学(trans-omics)”网络,描绘能量感知分子通过酶与信号蛋白向整个代谢通路传递信号的路径。
健康肌肉在压力下仍保持灵活
在瘦小鼠中,禁食引发了分子图景上广泛且高度协调的变化。关键的能量载体分子如 ATP 和 AMP 呈相反方向变化,导致 AMP/ATP 比率上升——这是能量匮乏的化学信号。这进一步强烈激活了 AMPK 通路,AMPK 是促进燃料消耗并抑制耗能过程的核心能量感应装置。许多酶在数量或磷酸化状态上随时间变化,蛋白质周转和脂肪利用等整体通路协同调整。作者将这种分子随时间起伏的模式称为“反应性”,在健康的肌肉和肝脏中这种反应性是普遍存在的。
肥胖肌肉停留在“差异”而非“响应”上
在肥胖的 ob/ob 小鼠中,情况明显不同。在所有分子层面上,禁食期间显示出真实时间过程变化的代谢物和蛋白质要少得多。尤其在蛋白质水平上的应答在肥胖肌肉中几乎缺失。取而代之的是大量酶(尤其参与脂肪代谢的酶)在肥胖动物中持续偏高,无论禁食时间如何,都表现为慢性升高。研究者将此称为“差异”:肥胖组织与健康组织之间的静态不一致,而不是随能量状况动态跟随的改变。换言之,肥胖组织进入禁食时已处于改变状态,然后几乎不再调整,而不是像健康组织那样灵活应对条件变化。
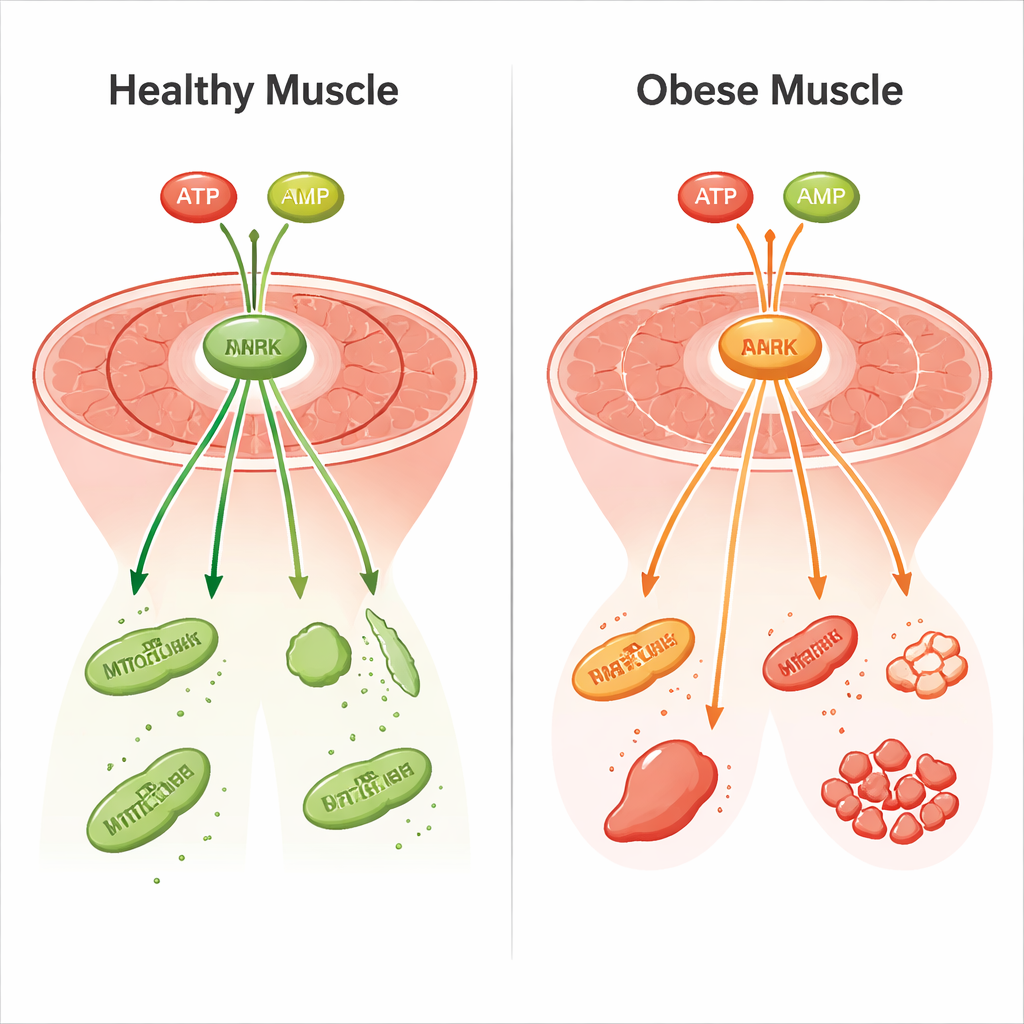
能量感应器在肥胖中沉默
聚焦能量感应回路揭示了一个关键故障。在瘦小鼠中,禁食提高了 AMP 并降低了 ATP,提升了 AMP/ATP 比率,并通过磷酸化显著激活 AMPK。下游变化与增强脂肪燃烧、降低脂质合成以及在肌肉和肝脏中减少糖原(储存的碳水化合物)合成相一致。在肥胖小鼠中,AMP 和 ATP 几乎没有变化,AMPK 激活被削弱或丧失,下游酶也没有按高效燃料切换所期望的方式发生转变。与此同时,蛋白质合成的标志物倾向于维持较高水平、蛋白质分解的标志物相对较低,这有助于解释为何肥胖组织含有更多的酶蛋白却缺乏协调的快速调控。
无法适应的机体
通过并列比较骨骼肌和肝脏,研究显示这一模式具有系统性:在肥胖中,两种器官都表现出对禁食反应性的全面丧失以及酶水平持续升高的倾向。肥胖体内并不像一台聪明、可调的发动机,而更像一个旋钮被卡住的机器——某些部位输出偏高,但在环境变化时难以调整燃料选择和能量使用。对非专业读者来说,结论是:肥胖不仅仅是“多余的脂肪”;它是对正常帮助我们应对食物短缺的紧急系统的一种全身性削弱,这种被削弱的灵活性可能是许多代谢并发症的根源。
引用: Li, D., Morita, K., Kokaji, T. et al. Global loss of metabolic responsiveness and elevated enzyme in leptin deficient obese mice during starvation. npj Syst Biol Appl 12, 53 (2026). https://doi.org/10.1038/s41540-026-00678-3
关键词: 代谢柔韧性, 禁食与饥饿, 肥胖与能量代谢, 骨骼肌与肝脏, AMPK 信号通路