Clear Sky Science · zh
PsAvh109通过模仿TPL功能抑制SA诱导的免疫,破坏Mediator复合体组装
一种作物杀手如何智胜植物防御
植物无法逃避病害,只能依赖写入基因组中的精细免疫系统。本研究揭示了一种主要的大豆病原如何窃听植物自身的警报信号,然后直击其基因控制中心,在关键防御基因应当被激活之时将其关闭。理解这种分子伏击为培育或改造能在毁灭性病害面前占据先机的作物提供了新线索。
植物的警报信号及其权衡
当微生物入侵时,植物会迅速重编程数千个基因,从生长转向防御。一个核心警报分子水杨酸(SA)会在受攻击的组织中激增,帮助激活所谓的病程相关(pathogenesis-related)基因,从而阻止嗜营养性病原,如许多真菌和卵菌。但这种强烈反应也伴随代价:用于防御的资源无法用于生长。为维持平衡,植物依赖灵活的遗传开关,能够根据不断变化的信号迅速关闭或开启防御基因。
细胞中免疫基因的“混音台”
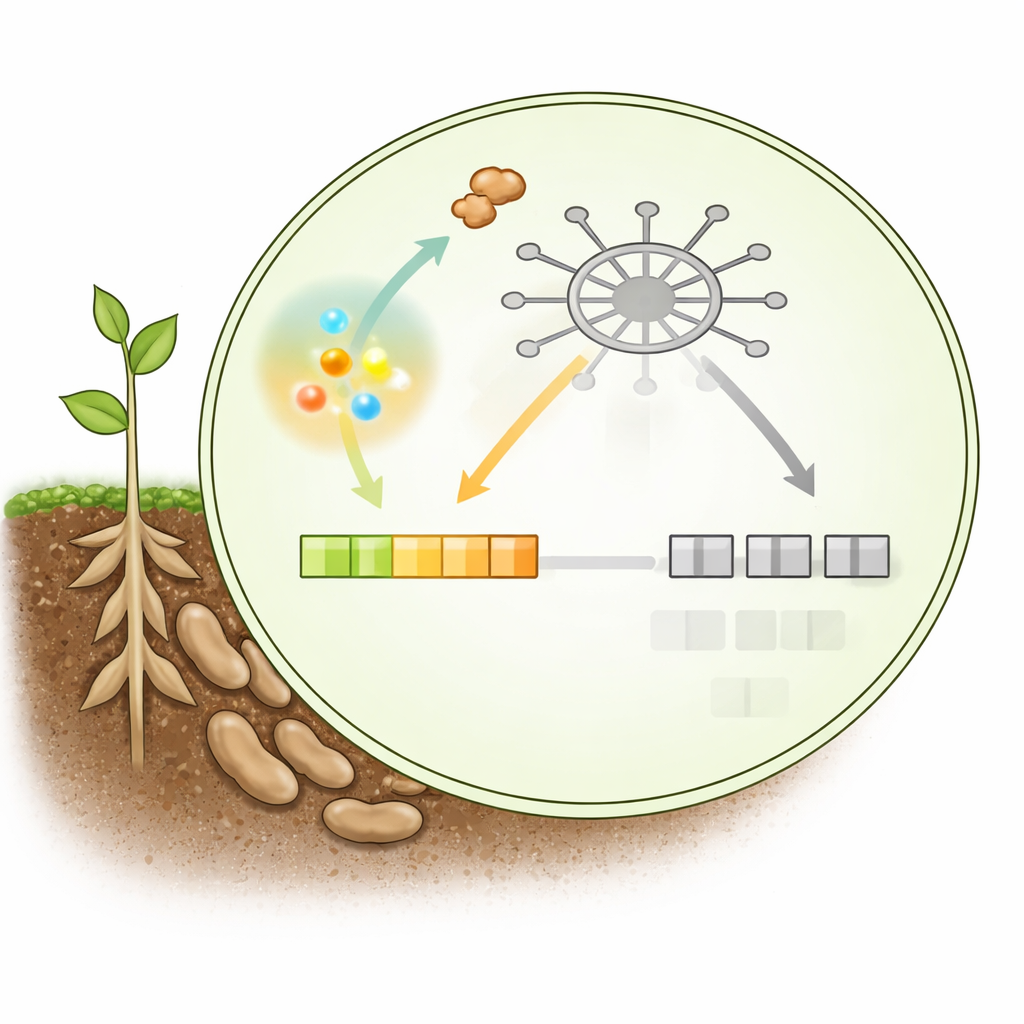
这一开关系统的核心是Mediator复合体,这是一个多蛋白枢纽,将结合DNA的调控因子与负责转录的酶连接起来。其两个组分,称为MED21和MED6,形成了一个关键接触点:当它们连接时,有助于组装激活SA响应防御基因所需的转录机器。植物还使用一种内置的刹车蛋白TOPLESS(TPL),它通常结合MED21的相同区域,阻止MED6对接。在平稳条件下,这可抑制免疫以便维持生长。
模仿植物自身刹车的病原效应子
大豆病原卵菌Phytophthora sojae产生一种名为PsAvh109的核定位蛋白,它被发现是该系统的主导破坏者。研究者发现,宿主来源的SA在培养条件和真实感染过程中都能强烈诱导病原体中PsAvh109的表达。被传入植物细胞后,PsAvh109进入细胞核并结合MED21,且正是结合在通常供MED6和TPL占据的小的N端区域。通过占据此位点,PsAvh109阻断MED21–MED6配对,妨碍在SA响应启动子上正确组装转录机器,包括若干PR1防御基因。在感染缺失PsAvh109的病原时,植物这些基因在早期的激活明显增强且病害减轻,而被强制定位至细胞质的PsAvh109变体则失去促进感染的能力。
劫持植物自身信号的时序
该研究揭示了一个引人注目的反转:本应释放植物自有刹车的同一SA爆发反而触发了病原体的反击。在健康状态下,TPL位于MED21上并抑制防御基因活性。当感染时SA上升,TPL与MED21的结合被削弱,允许MED6对接并打开免疫基因。但PsAvh109在病原体内也会被SA诱导,其与MED21的结合甚至比MED6或TPL更牢固。它在TPL被置换时进入植物细胞核,快速重新占据MED21,使复合体在有SA的情况下仍然保持被抑制状态。来自相关Phytophthora物种的同源效应子也显示出类似的SA诱导性,提示这种“感知并回应”的策略在广泛物种中保守存在。
对作物保护的启示
通过描绘这场分子博弈,作者揭示了病原体利用宿主信号节律的新途径:它们不仅仅阻断识别,而是在免疫被激活时精准时机释放效应子并直接重塑基因控制。对非专业读者而言,关键的信息是抗病性不仅依赖于是否具备合适的防御基因,还取决于谁控制着操作这些基因的开关。针对PsAvh109–MED21相互作用、增强MED21–MED6结合或改变MED21接触表面,可能为培育在聪明病原面前仍能维持强烈SA触发免疫的大豆及其他作物提供新途径。
分子伏击的可视化快照
概括而言,这项工作描绘了植物细胞核内单一对接位点上的三方竞争:偏向生长的抑制因子、激活防御的配对因子以及病原效应子共同争夺控制权。在植物处于静止时,抑制因子占据该位点并将免疫基因维持在低水平;当SA激增时,植物短暂占优,抑制因子被置换,激活因子得以开启防御基因。但病原体感知到这一SA激增后迅速部署PsAvh109,其以更高亲和力抓住相同位点,将那些基因再次逼回沉默状态。
引用: Tan, X., Sun, Y., Qi, Z. et al. PsAvh109 suppresses SA-triggered immunity by mimicking TPL function to disrupt mediator complex assembly. Nat Commun 17, 3486 (2026). https://doi.org/10.1038/s41467-026-71909-8
关键词: 植物免疫, 水杨酸信号传导, 大豆疫霉(Phytophthora sojae), 转录抑制, Mediator复合体