Clear Sky Science · zh
在无端粒酶细胞中,最短端粒既引发基因组不稳定又导致复制性衰老
为什么微小的染色体帽对衰老和癌症很重要
每次细胞分裂时,染色体末端的保护帽——端粒——都会逐渐变短。当端粒过短时,细胞停止分裂,这有助于抑制肿瘤,但也与衰老相关。本文在面包酵母中提出了一个简单却重要的问题:是否仅仅是一个异常短的端粒就能使细胞停下来,而这个脆弱点是否也是危险DNA重排开始的部位?
一种制造精确定长短端粒的新方法
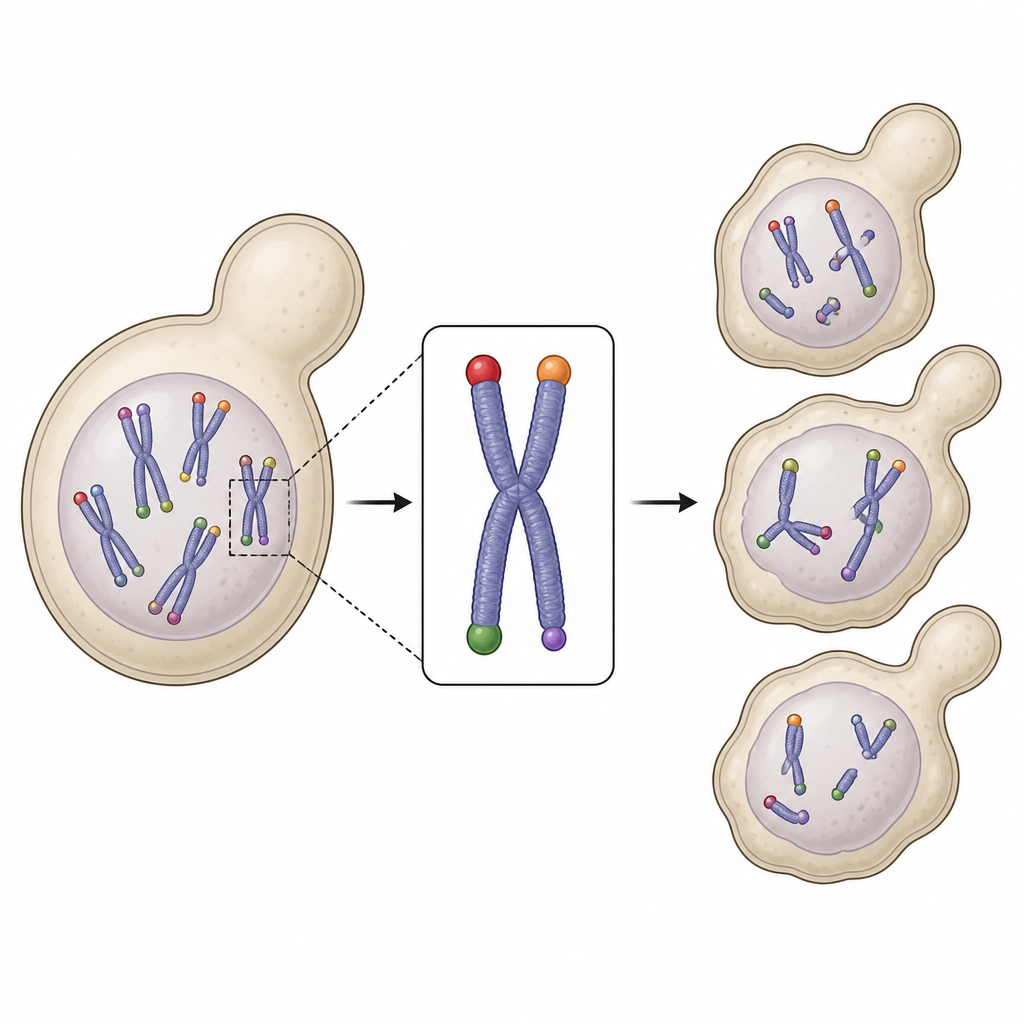
为了解答这个问题,研究者在酵母中构建了一个称为 FinalCut 的遗传工具。它利用可编程的DNA切割酶,将所选的某一个染色体末端修剪到精确长度,同时保持其他端粒不变。他们还可以开关细胞的天然端粒延长酶端粒酶。这样就可以观察单个受控短端粒随时间的行为,而这在天然存在的不同长度端粒混杂中通常是看不清的。
观察细胞停止分裂前的最后步骤
将 FinalCut 与单分子DNA测序结合后,团队追踪了工程化短端粒随每次细胞分裂的缩短过程。他们发现端粒每轮仅丢失少量的DNA“字母”,但当该工程化端粒缩短到大约30到40个碱基时,其行为就发生了显著变化。该染色体末端不再被维持,开始向内侵蚀。与此同时,在微流控小室中的单细胞追踪显示,酵母谱系会按由该单个端粒初始长度决定的世代数正常分裂,然后突然进入周期显著延长、应激的细胞周期阶段并最终死亡。结合这些数据的数学建模指向一个清晰阈值:当最短端粒低于临界大小时,它会可靠地把细胞置于永久停滞状态。
基因组混乱真正的起点在哪里
作者随后探问在端粒变短时,严重DNA损伤从何而来。他们在工程化端粒附近和远端的染色体末端分别安置了遗传“报告器”片段。当关闭端粒酶并允许工程化末端缩短时,最短端粒附近的突变率急剧上升,而远处区域则相对平静。其中大多数变化不是简单的碱基错误,而是大规模的重排——靠近脆弱端粒的染色体片段与另一条染色体臂发生融合。
从磨损的帽子到危险重排的逐步路径
对突变酵母进行全基因组测序证实了一个明确的模式。受损的染色体末端倾向于连接到共享附近序列的其他染色体末端,产生一种称为非互惠易位的单向融合。这些事件依赖于一种特定的DNA复制因子 Pol32,该因子对一种可从模板复制长片段DNA的修复过程是必需的。实质上,一旦最短端粒跌破保护阈值,末端就被咬回,暴露出可配对的序列,这些序列随后劫持该修复通路,从基因组的其他部位重建出一个新的、更长的染色体末端。
这对衰老细胞和肿瘤意味着什么
综合所有证据,研究表明在无端粒酶的酵母细胞中,单个最短端粒既决定细胞何时停止分裂,又标记了危险DNA重排开始的热点。随着细胞老化并且该端粒越过关键最小长度,它既触发DNA损伤警报以停止生长,又增加了局部重排的可能性,有时通过重建断裂末端使细胞暂时逃脱停滞。由于端粒生物学和修复通路高度保守,这些结果为人类细胞中某个尤其短的染色体末端如何同时限制细胞分裂次数并在相同条件下激发有助于癌变的基因组变化提供了具体图景。
引用: Berardi, P., Martinez-Fernandez, V., Rat, A. et al. Both genome instability and replicative senescence stem from the shortest telomere in telomerase-negative cells. Nat Commun 17, 4271 (2026). https://doi.org/10.1038/s41467-026-70352-z
关键词: 端粒, 细胞衰老, 基因组不稳定性, 端粒酶, 酵母模型