Clear Sky Science · zh
双联微管相关的tektin和酶差异性调控精子鞭毛的完整性与运动性
为何微小的尾部结构会影响生育力
对精子来说,游动能力至关重要。每个精子依靠一条细长的、鞭状的尾巴推动其到达卵子;当这条尾巴构建不正确或摆动方式异常时,就可能导致不育。本研究深入尾部的显微支架,探讨一个看似简单但重要的问题:哪些特定分子维持精子尾部的完整并确保正常运动,哪些分子在内部悄然调节其摆动?回答这一问题有助于解释男性中不同类型的精子活动力差异,并指向更精确的男性不育诊断方向。
精子尾部的内部骨架
每条精子尾部的核心是一种高度有序的支架,由空心的管状结构以经典的“9+2”模式排列:九对外周管围绕着两根中央管。这些双联管并非空无一物;它们内衬并装饰着数十种专门蛋白,这些蛋白加固结构并协调其摆动。作者聚焦于小鼠精子中的四种此类蛋白:两种形成丝状的“tektin”,用于加固管腔内部,和两种附着在相同框架上的酶——一类激酶和一类磷酸酶。通过高分辨率冷冻电子显微镜,他们绘制了这些蛋白的定位,并显示与呼吸道等类似摆动结构相比,精子尾部在结构上更为复杂,具有专为生殖调优的额外成分。 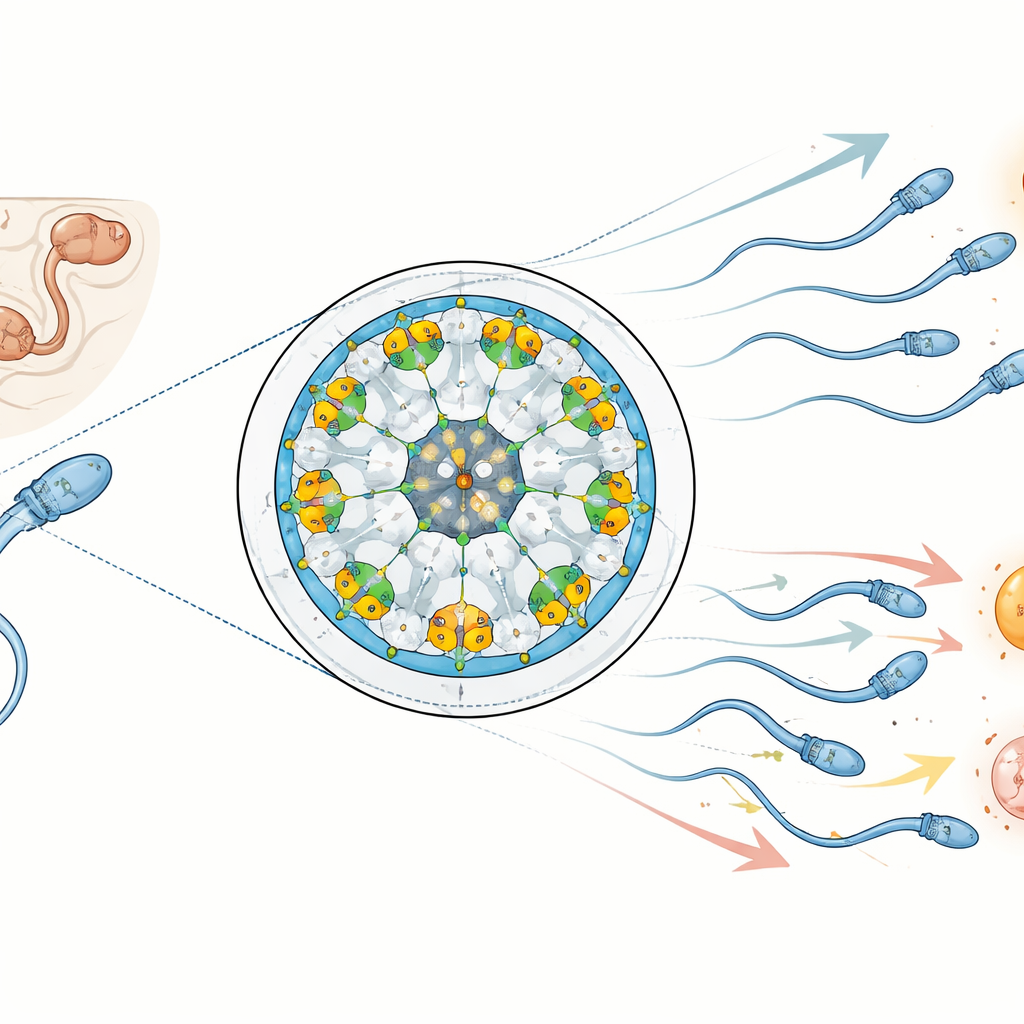
在小鼠中关闭关键基因
为了解这些蛋白在活体中的实际功能,研究团队构建了四条敲除小鼠系,分别完全缺失其中一个目标基因:Tekt1、Tekt5、Tssk6或Dusp21。随后他们检查了生育能力、精子计数、尾部形态和游动能力。缺失Tekt1或Tssk6的雄性尽管睾丸外观正常且精子数量正常,但完全不育。相比之下,缺失Tekt5或Dusp21的雄性能产生与健康小鼠相似的后代,尽管在某些情况下尾部行为出现细微变化。该结果表明,并非所有看似相似的尾部蛋白重要性相同:有些对生育至关重要,另一些则更可有可无或可被其他分子补偿。
结构丝如何维持尾部稳定性
tektin像沿着双联管内部延伸的加固杆。在缺失TEKT1的精子中,冷冻电子显微镜显示整个内部tektin丝束几乎完全缺失,同时数种依赖该丝束锚定的伴随蛋白也消失。总体的管状排列仍可见,但更常出现错位或部分紊乱,尾部更易弯曲或盘绕。使用超声波等机械试验使结构震散的实验表明,这些缺乏tektin的尾部更容易分解为自由构件,显示支架更脆弱、强度更低。相比之下,缺失精子特异性TEKT5的小鼠仅丢失一小部分内部蛋白,对尾部结构和游动性的影响较轻,这进一步支持了保守的TEKT1为运动的主干束这一观点。
内部酶如何微调运动
研究的两种酶位于或靠近同一双联管,但以不同方式工作:它们通过添加或移除磷酸基团来调节许多尾部蛋白的化学“开/关”状态。激酶TSSK6被证明至关重要。来自Tssk6敲除小鼠的精子常常完全缺失尾部或尾部严重弯曲,运动模式极为异常。对蛋白磷酸化模式的详细分析显示,许多尾部组分的数百个位点发生失调,包括那些对头部与尾部连接重要的位点。换言之,当这一单一酶缺失时,协调摆动和结构完整性的整个内部信号网络都会紊乱。相比之下,移除磷酸酶DUSP21产生的精子出乎意料地接近正常,提示其他磷酸酶可补偿其缺失。
将小鼠遗传学与人类不育联系起来
通过将深度结构成像与全局蛋白及磷酸化测量相结合,研究表明一些内部尾部蛋白主要充当物理加固件,而另一些则作为摆动机械的控制旋钮。缺失TEKT1会导致看起来大体正常但游动乏力的精子——这一模式类似于某些患者,他们的精子尾部结构完整却活动迟缓。另一方面,缺失TSSK6则引起剧烈的尾部畸形,类似于男性中一种严重病症,其中许多精子缺失、弯曲或盘绕尾部。 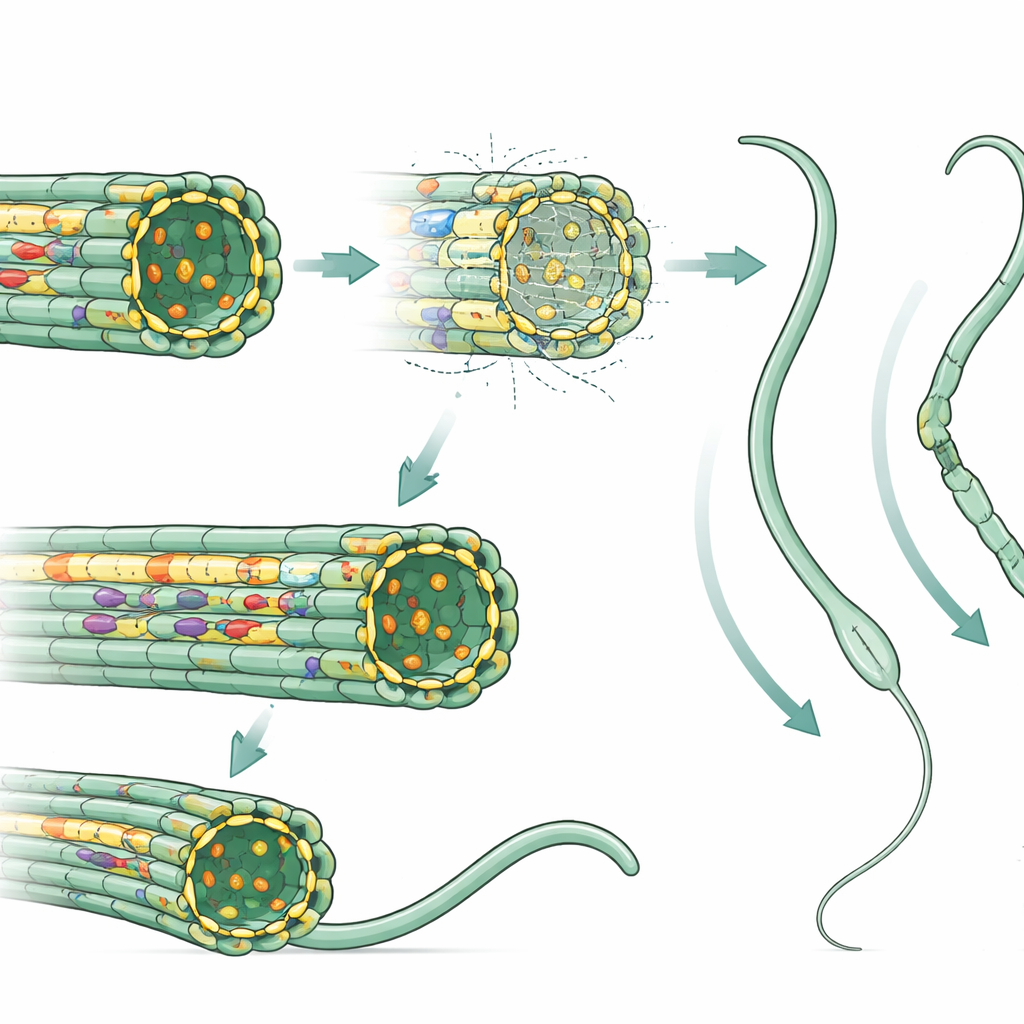
引用: Liu, Q., Zhou, L., Liang, X. et al. Doublet microtubule-associated tektins and enzymes differentially regulate sperm flagellar integrity and motility. Nat Commun 17, 3316 (2026). https://doi.org/10.1038/s41467-026-69714-4
关键词: 精子鞭毛, 男性不育, 微管结构, 蛋白质磷酸化, 小鼠遗传学