Clear Sky Science · ru
iAODE для бенчмаркинга и континуального моделирования одно-клеточной доступности хроматина
Проследить путь отдельных клеток
Наши тела состоят из триллионов клеток, которые непрерывно меняются по мере роста, заживления и старения. Современные методы одно-клеточного секвенирования позволяют «заморозить» эти клетки в момент времени и узнать, какие участки ДНК раскрыты и активны. Но такие снимки данных шумны и фрагментарны, что затрудняет отслеживание непрерывных путей, по которым клетки проходят в процессе созревания или смены судьбы. В этой статье представлен новый вычислительный фреймворк iAODE, который превращает эти разрозненные кадры в гладкие «фильмы» клеточных изменений и предлагает строгий способ оценить, насколько хорошо разные методы восстанавливают такие клеточные истории.
Почему важна открытость хроматина
Гены включаются и выключаются не только по их последовательности, но и по тому, открыт ли окружающий ДНК фрагмент и доступен ли он. Метод single-cell ATAC-seq измеряет эти открытые_regions по отдельным клеткам. Однако полученные данные крайне разрежены: большинство позиций выглядят как нули — либо потому, что ДНК действительно закрыта, либо потому, что она просто не была захвачена. Существующие инструменты обычно сжимают такие данные в низкоразмерное представление, а затем, как отдельный шаг, пытаются восстановить развитие, упорядочивая клетки по псевдовремени. Это разделение означает, что модели не обучаются явно учитывать плавное временное изменение, и неясно, какие типы внутренних представлений лучше всего восстанавливают реальные биологические траектории.
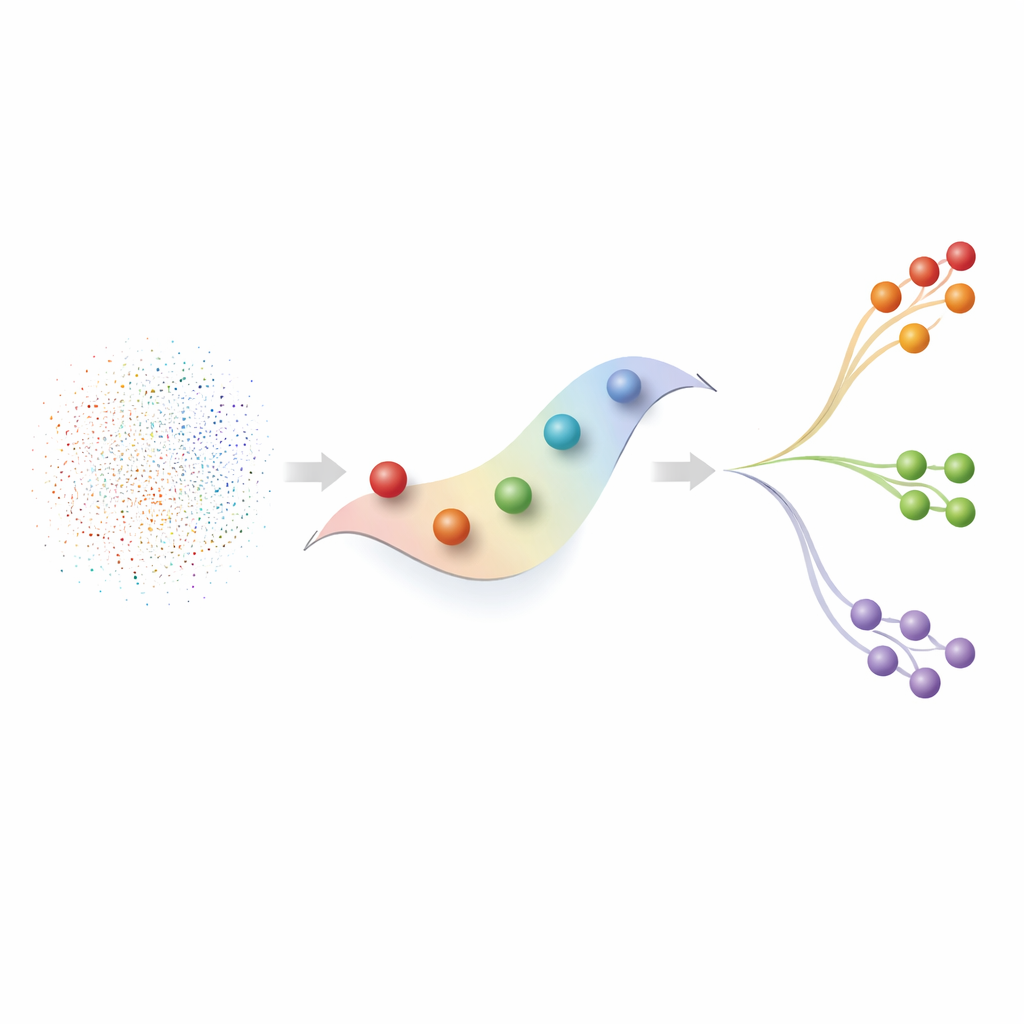
Новый способ изучать непрерывные клеточные пути
iAODE переосмысливает этот конвейер, встроив непрерывность в ядро модели. Он стартует с популярного фреймворка глубокого обучения (вариационного автоэнкодера), который учится компактному «латентному» пространству, улавливающему ключевые шаблоны в данных. Поверх него авторы добавляют нейронное обыкновенное дифференциальное уравнение (Neural ODE), которое рассматривает положение каждой клетки в латентном пространстве как точку на гладкой временной траектории. Кодировщик не только сжимает данные, но и предсказывает относительное время для каждой клетки, в то время как компонент ODE учится тому, как латентные состояния меняются от ранних к поздним позициям. Два дополнительных решения — ослабление стандартного штрафа, заставляющего латентные измерения вести себя независимо, и добавление сжатого «бутылочного горлышка» для восстановления — поощряют совместное изменение групп признаков в биологически значимые модули и помогают отфильтровывать шум.
Создание честной площадки для сравнения
Чтобы оценить iAODE и отдельные элементы его дизайна, авторы собрали одну из крупнейших стандартизированных коллекций бенчмарков в этой области: 248 наборов данных по одно-клеточной доступности хроматина и 123 набора одно-клеточной РНК, охватывающие разные виды, технологии и размеры. Они также разработали набор из 20 метрик, которые отдельно измеряют три аспекта: насколько непрерывными и правильной формы являются латентные траектории, насколько достоверно сохраняется структура при проекции для визуализации и насколько хорошо захватываются кластеры и регуляторные связи. Сначала протестировав эти метрики на тщательно смоделированных данных с известными траекториями, авторы показали, что оценки меняются плавно и предсказуемо при увеличении или уменьшении непрерывности — важная проверка здравого смысла перед применением к реальной биологии.
Что показывают бенчмарки
Вооружившись этой площадкой, команда систематически анализирует iAODE. Включение и выключение отдельных компонентов показывает, что компонент ODE в основном обеспечивает глобальный каркас и чёткую направленность изменений, тогда как низкопенализирующая регуляризация и бутылочное горлышко уточняют геометрию и делают модель более устойчивой к шуму. На сотнях наборов данных по хроматину полная модель iAODE последовательно даёт более плавные и более стойкие траектории по сравнению с установленными глубокими генеративными методами, такими как scVI, PeakVI, и более ранним инструментом на основе ODE, а также по сравнению с традиционными методиками, такими как метод главных компонент и diffusion maps. Та же картина наблюдается при применении фреймворка к данным одно-клеточной РНК, что указывает на то, что основные идеи обобщаются за пределы хроматина. Визуальные примеры из наборов данных мозга и крови показывают, что латентные оси iAODE согласуются с известными маркерными генами и регуляторными регионами, а группы латентных признаков обогащены ожидаемыми биологическими процессами, что указывает на то, что изученные траектории не только плавные, но и интерпретируемые.

Почему это важно для понимания судьбы клеток
По сути, эта работа утверждает, что непрерывность в клеточном развитии следует считать принципом проектирования, а не побочным эффектом. Встроив гладкую динамическую модель в шаг представления и тщательно ослабив ограничения, которые в противном случае разрушили бы скоординированные сигналы, iAODE создаёт латентные пространства, где состояния клеток формируют текущие пути, а не разрозненные кластеры. Сопровождающие наборы данных бенчмарков и метрики дают сообществу общую мерку для сравнения методов, нацеленных на восстановление таких путей. Для исследователей это значит более надёжные карты того, как клетки проходят развитие, иммунные ответы или болезни, и более ясный способ связать эти пути с лежащими в основе регуляторными программами, закодированными в геноме.
Цитирование: Fu, Z., Chen, C., Wang, S. et al. iAODE for benchmarking and continuum modeling of single-cell chromatin accessibility. Commun Biol 9, 507 (2026). https://doi.org/10.1038/s42003-026-09768-8
Ключевые слова: одно-клеточная геномика, доступность хроматина, моделирование траекторий клеток, глубокие генеративные модели, Neural ODE