Clear Sky Science · ru
Ацетилхолин разделяет смешанные сигналы дофамина, влияющие на обучение и движение
Почему это важно для повседневного поведения
Каждый раз, когда вы узнаёте, что звук означает «лакомство будет» или чувствуете прилив энергии, чтобы двигаться быстрее, в вашем мозге работают крошечные химические сигналы. Два из самых значимых — дофамин, часто ассоциируемый с наградой, и ацетилхолин, менее известный, но мощный модулятор. В этом исследовании показано, что важно не только количество высвобождаемого вещества, но и точный момент их появления относительно друг друга — синхронизация, которая может определить, будете ли вы учиться на опыте или просто двигаться быстрее.
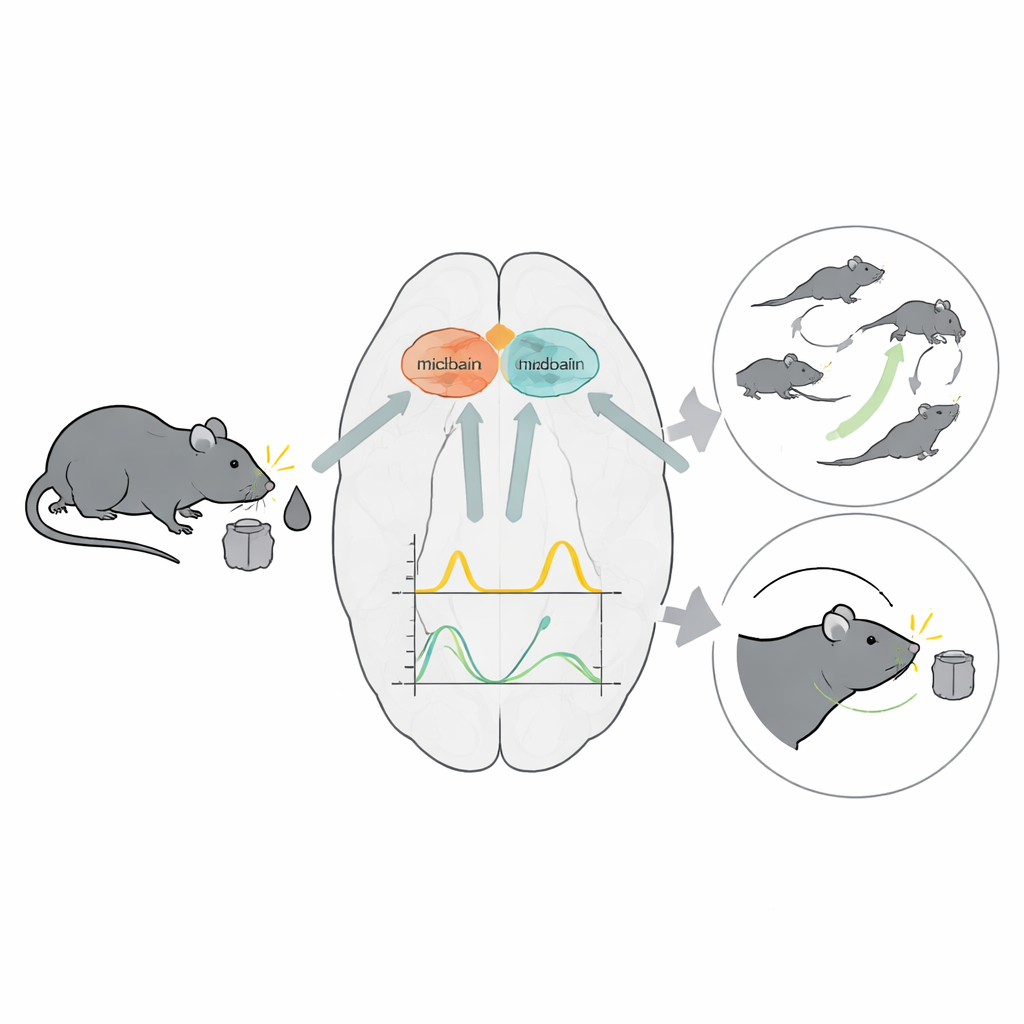
Два посредника мозга с разными задачами
Нейроны, синтезирующие дофамин, в глубине среднего мозга посылают широко разветвлённые волокна в стриатум — область мозга, ключевую для обучения действиям, ведущим к награде, и для контроля движений. На протяжении многих лет исследователи знали, что дофамин может одновременно обучать животных тому, какие выборы ценны, и усиливать их движения. Загадкой оставалось, как один и тот же химический сигнал может нести и информацию об обучении, и сигналы, связанные с движением, не смешиваясь для принимающих нейронов. Подозревали, что ацетилхолин, высвобождаемый редким классом стриатальных клеток — холинергическими интернейронами, помогает сортировать или «размешивать» эти перекрывающиеся сообщения, но эта идея не была строго проверена во время реального поведения.
Задача, разделяющая обучение и движение
Чтобы разобраться, исследователи обучили крыс самостоятельно выполнять задачу «временного торга», которая чётко отделяла события, связанные с наградой, от событий, связанных с движением. В каждом испытании звук оповещал о размере предлагаемой воды; позже свет указывал, в каком боковом порте она может появиться, после непредсказуемой паузы. Крысы могли либо продолжать ждать, либо бросить испытание и начать новое, что фактически показывало, как они ценят текущее предложение по сравнению с будущими. Такой дизайн создавал моменты, когда животное обновляло свои ожидания по поводу награды, и другие моменты, когда оно делало быстрые ориентировочные движения головой, что позволяло ученым сравнить сигналы дофамина и ацетилхолина в этих разных контекстах.
Как синхронизация решает: обучение или скорость
Используя светочувствительные датчики, команда измеряла быстрые изменения дофамина и ацетилхолина в дорсомедиальном стриатуме во время выполнения задания. Когда звуки впервые объявляли размер возможной награды, дофамин показывал кратковременные всплески, соответствующие классическим сигналам «ошибки прогноза» — разнице между ожидаемым и полученным. В те же моменты ацетилхолин снижался, причём этот спад чуть предшествовал всплеску дофамина. При такой последовательности больших всплесков дофамина предсказывали, как крысы скорректируют поведение в следующем испытании, например начнут ли они быстрее запускать новый пробег после недавней награды. Нейроны, регистрируемые с помощью тонких электродов, меняли шаблоны своей активности от испытания к испытанию в виде, согласующемся с длительной синаптической пластичностью, что указывает на то, что эти всплески дофамина, приходящие вскоре после пауз ацетилхолина, приводили к изменениям, связанным с обучением в цепи.
Когда тот же дофамин перестаёт обучать
Картина менялась при другом ключевом событии: когда период ожидания заканчивался и награда становилась доступной после короткой или длинной задержки. Здесь всплески дофамина снова отражали ошибки прогноза — они были больше при необычно длинной задержке — но теперь они возникали чуть раньше, а не после спадов ацетилхолина. Несмотря на формальное сходство с учебными сигналами, эти всплески дофамина не предсказывали никаких измеримых изменений в последующем поведении крыс. Животные не стали систематически дольше ждать, быстрее тыкать или изменять время начала испытаний после длинных задержек. Иными словами, тот же тип дофаминового сигнала, сдвинутый немного раньше относительно ацетилхолина, уже не вызывал наблюдаемого обучения.
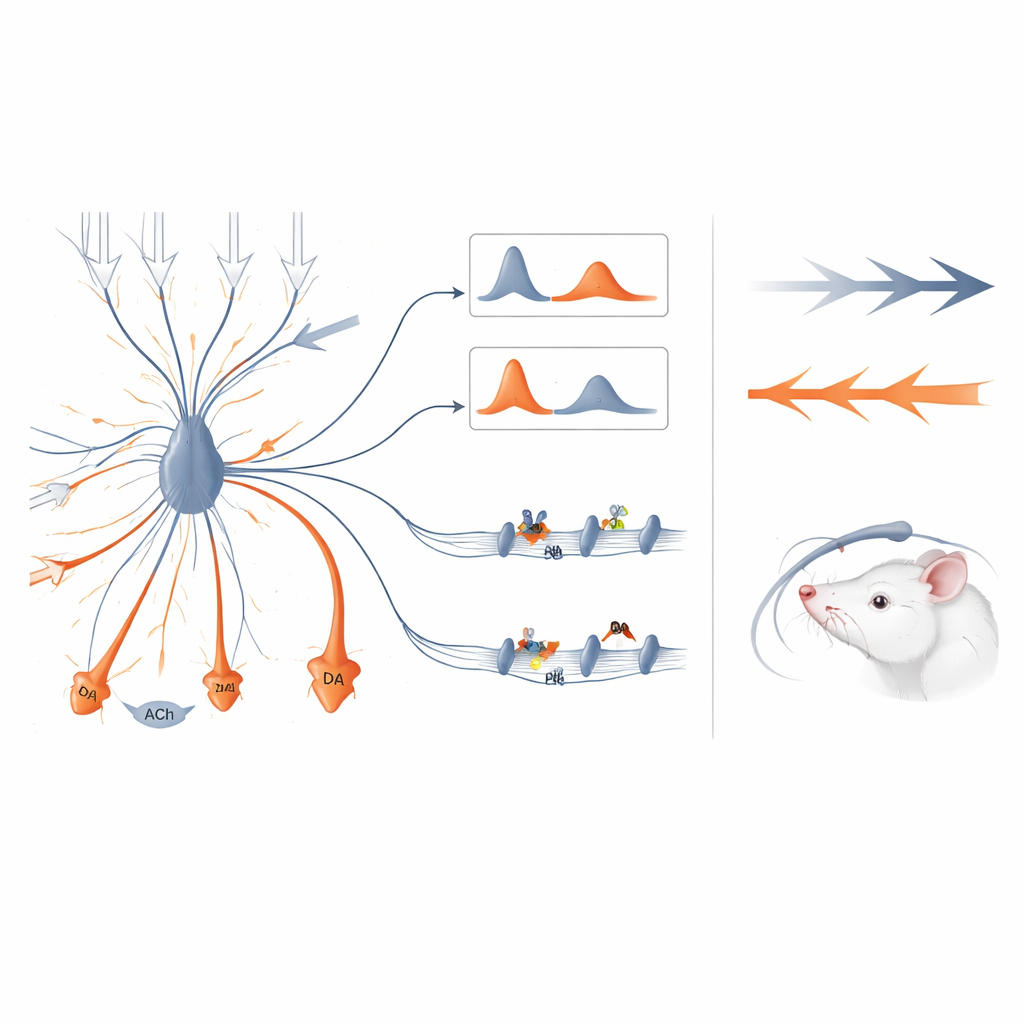
Переход от обучения к усилению движения
Иная картина появилась в моментах, доминируемых движением. Когда зажигался боковой свет и крыса резко поворачивала голову к потенциальному порту с наградой, дофаминовые сигналы в стриатуме были сильнейшими для движений в сторону, противоположную месту регистрации, и усиливались по мере увеличения скорости ориентировочного движения. В такие моменты ацетилхолин не снижался; он наоборот проявлял всплеск почти синхронно с дофамином. Сила дофаминового сигнала предсказывала, насколько энергичным будет предстоящий двигательный акт, но не оставляла того долговременного следа в активности нейронов, который наблюдали при событиях, связанных с обучением. По сути, когда дофамин и ацетилхолин росли вместе, дофамин скорее выступал как сигнал «двигайся быстрее», а не «обнови свои ожидания».
Что это значит для обучения, движения и болезней
В сумме результаты говорят о том, что ацетилхолин действует как временной шлюз для влияния дофамина. Когда ацетилхолин кратко приостанавливается, а дофамин следует вскоре после этого, дофамин наиболее эффективен в перестройке связей в стриатуме и поддерживает обучение тому, какие действия ценны. Когда дофамин предшествует или совпадает с всплесками ацетилхолина, тот же химический сигнал скорее направляется не на долговременные изменения, а на энергизацию текущих движений. Эта тонкая временная регуляция помогает мозгу не смешивать учебные и моторные сигналы и даёт новые представления о расстройствах, таких как болезнь Паркинсона, при которых нарушены обе системы — дофаминергическая и холинергическая.
Цитирование: Jang, H.J., McMahon Ward, R., Golden, C.E.M. et al. Acetylcholine demixes heterogeneous dopamine signals for learning and moving. Nat Neurosci 29, 840–850 (2026). https://doi.org/10.1038/s41593-026-02227-x
Ключевые слова: дофамин, ацетилхолин, обучение с подкреплением, пластичность стриатума, энергичность движений