Clear Sky Science · ru
Фруктозо-1,6-бисфосфат связывает гликолитическую активность и клеточную адгезию
Как сахар разговаривает с силой клеточного захвата
Каждый раз, когда клетка ползёт по поверхности, заживает рану или начинает разрастаться как опухоль, ей нужно решить, как крепко держаться и насколько решительно продвигаться вперёд. В этом исследовании показано, что знакомый продукт распада сахара внутри клетки — фруктозо-1,6-бисфосфат (ФБФ) — ведёт себя как молекулярный регулятор, который связывает скорость сжигания сахара клеткой с её способностью прикрепляться и распространяться. Понимание этого скрытого диалога между метаболизмом и клеточной адгезией может изменить наше представление о развитии, иммунитете и инвазии рака.
Открытие скрытой связи
Чтобы выяснить, что контролирует крошечные анкеры, которыми клетки хватаются за окружение, исследователи провели масштабный скрининг с инактивированием генов. Они использовали автоматическую микроскопию для проверки более 18 000 генов в человеческих клетках, отслеживая изменения «фокальных адгезий» — маленьких точечных структур, которые соединяют внутренний каркас клетки с внешним миром. Среди сильнейших попаданий оказался неожиданный участник: альдолаза A, классический фермент гликолиза, пути расщепления глюкозы для получения энергии. При снижении уровня альдолазы A клетки образовывали значительно больше фокальных адгезий и распространялись на большую площадь, и этот эффект исчезал при восстановлении нормальной версии фермента.
Метаболит сахара как сигнал, а не только топливо
На первый взгляд можно было бы предположить, что эффект связан просто с энергией: изменить гликолиз — изменить уровни АТФ — и поведение клеток сменится. Но выяснилось, что история тоньше. Когда команда снижала активность других ферментов гликолиза, во всех случаях падал уровень АТФ, однако только манипуляции с этапами, контролирующими уровни ФБФ, меняли адгезию. Снижение фермента PFK, который синтезирует ФБФ, приводило к сжатию клеток и утрате адгезий, что противоположно эффекту потери альдолазы. Удаление фермента на более позднем этапе пути мало влияло на форму клетки и её сцепление. Прямые измерения показали, что высокие уровни ФБФ хорошо коррелировали с крупными, сильно прикреплёнными клетками, тогда как низкие уровни ФБФ соответствовали мелким, слабо прикреплённым клеткам. Блокирование производства ФБФ, генетически или с помощью ингибитора гликолиза, могло «вернуть» клетки с дефицитом альдолазы к норме, доказав, что именно концентрация этого метаболита — а не общие запасы энергии — определяет адгезию.
От внутренней химии к внешней форме
Как небольшой метаболит перестраивает внешний край клетки? При наблюдении в реальном времени маркеров фокальной адгезии снизу клетки учёные обнаружили, что ФБФ стимулирует появление новых адгезий, а не замедляет их распад. Высокий ФБФ увеличивал скорость сборки адгезий и число новых сайтов, тогда как низкий ФБФ действовал обратным образом. Одновременно перестраивался актиновый цитоскелет — динамическая сеть белковых филаментов, двигающая клетку. Клетки с высоким ФБФ демонстрировали широкие пластинчатые выросты и более сложные актиновые узоры, они быстрее расползались по поверхности, тогда как ФБФ‑бедные клетки показывали меньше стресс‑волокон и менее выраженный выступающий край. Важно, что естественные ситуации, требующие активного расползания и миграции — например, пересев или движение клеток — сопровождались сильными подъёмами ФБФ до уровней, сопоставимых с экспериментальными манипуляциями.
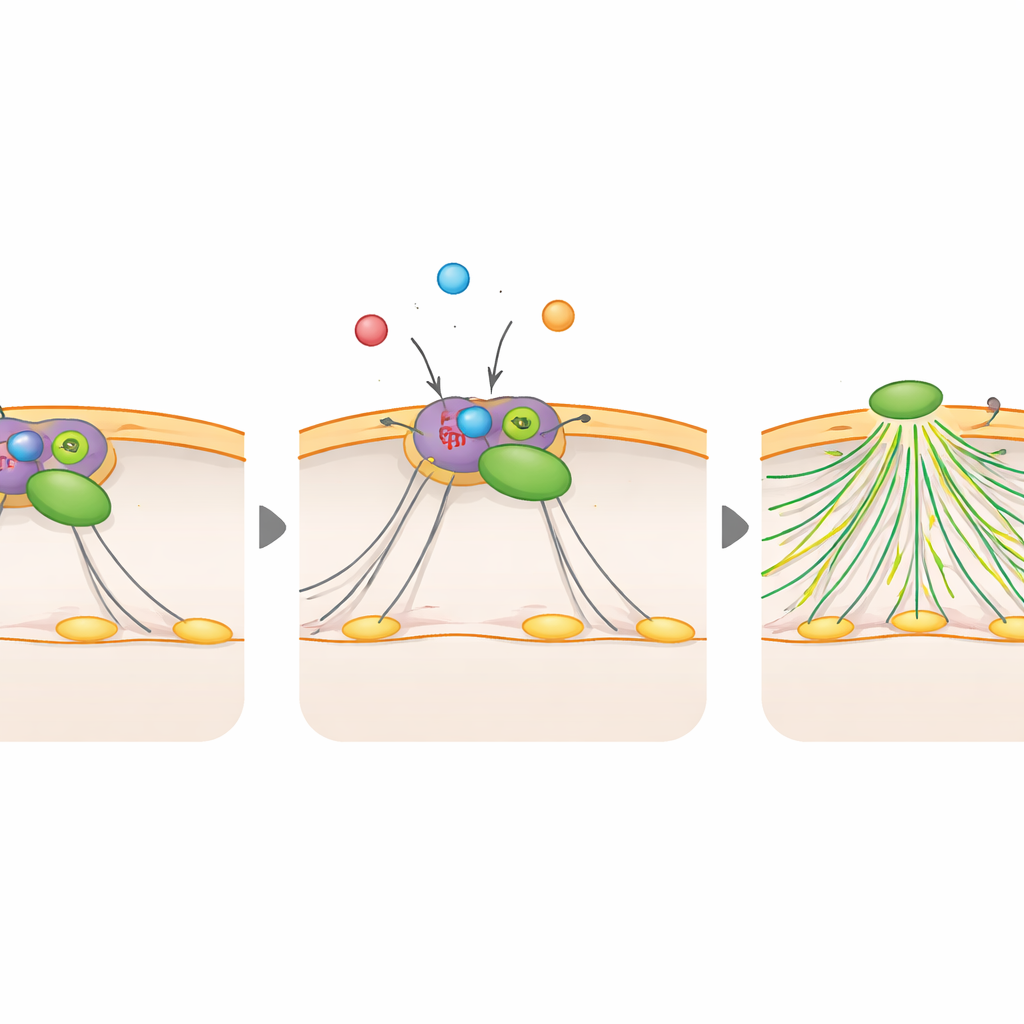
Ослабление молекулярного тормоза
Чтобы связать это поведение с известными сигнальными цепями, команда искала белки, которые меняют конформацию при контакте с ФБФ в клеточных экстрактах. Поиском был выявлен Rac1, главный регулятор актиновых выростов, и RCC2, белок, который может удерживать Rac1 в неактивном комплексе. Исследователи показали, что высокий ФБФ повышает активность Rac1, а низкий ФБФ её подавляет. При удалении Rac1 или его фиксации в неактивной форме дополнительные адгезии и распластывание, вызванные высоким ФБФ, исчезали; при запирании Rac1 в активном состоянии он обходил эффекты низкого ФБФ. Биохимические тесты продемонстрировали, что ФБФ связывается прямо с RCC2 и ослабляет его взаимодействие с Rac1. По сути, ФБФ вырывает Rac1 из-под его ингибитора, позволяя другим факторам активировать Rac1, что запускает перестройку актина, образование новых выростов и появление дополнительных сайтов адгезии.
Почему это важно для здоровья и болезни
Эта работа показывает, что ФБФ — не просто промежуточный продукт производства энергии, но и мессенджер, позволяющий клетке чувствовать, когда гликолиз активен, и корректировать своё физическое поведение. При усиленном расщеплении сахара ФБФ растёт, освобождает Rac1 от RCC2 и поощряет клетки к расползанию и исследованию; при низком гликолизе Rac1 остаётся сдержанным, и клетки остаются компактными и менее адгезивными. Такой механизм, вероятно, влияет на раннее развитие, где гликолиз особенно активен, а также на поведение раковых клеток, миграцию иммунных клеток и рост кровеносных сосудов — процессы, которые зависят от высокой гликолитической активности и динамичных клеточных выростов. Раскрывая, как простой метаболический интермедиат регулирует «захват» клетки за окружение, исследование подчёркивает прямую химическую связь между тем, что клетка «съела», и тем, как она движется.
Цитирование: Hoffmann, L., Duchmann, M., Lazarow, K. et al. Fructose-1,6-bisphosphate couples glycolytic activity to cell adhesion. Nat Cell Biol 28, 739–753 (2026). https://doi.org/10.1038/s41556-026-01911-1
Ключевые слова: клеточная адгезия, гликолиз, сигнализация Rac1, миграция клеток, инвазия рака