Как сперматозоиды защищают своё генетическое послание
Каждый здоровый сперматозоид должен передать потомству точную копию ДНК. Однако в процессе формирования половые хромосомы у мужчин, X и Y, сталкиваются с особой проблемой: они не совпадают так же ровно, как другие пары хромосом. В этом исследовании показано, как подвижная структура внутри клетки — ядрышко — временно перемещается к половым хромосомам и помогает выключить их гены на критическом этапе, защищая фертильность.
Зона тишины для половых хромосом
В яичках развивающиеся сперматоциты проходят длинную серию этапов, в ходе которых перестраивается и их ДНК, и клеточная архитектура. На ключевой стадии, называемой пахитеном, большинство хромосом тесно сопрягаются и обмениваются участками ДНК в подготовке к разделению на сперматозоиды. X и Y при этом имеют лишь небольшую общую область и в основном остаются несопоставленными. Чтобы избежать ошибок, клетка выключает большинство генов на X и Y в процессе, известном как мейотическая инактивация половых хромосом. Подавленные хромосомы образуют отдельную каплеобразную область у края ядра, называемую XY-телом.
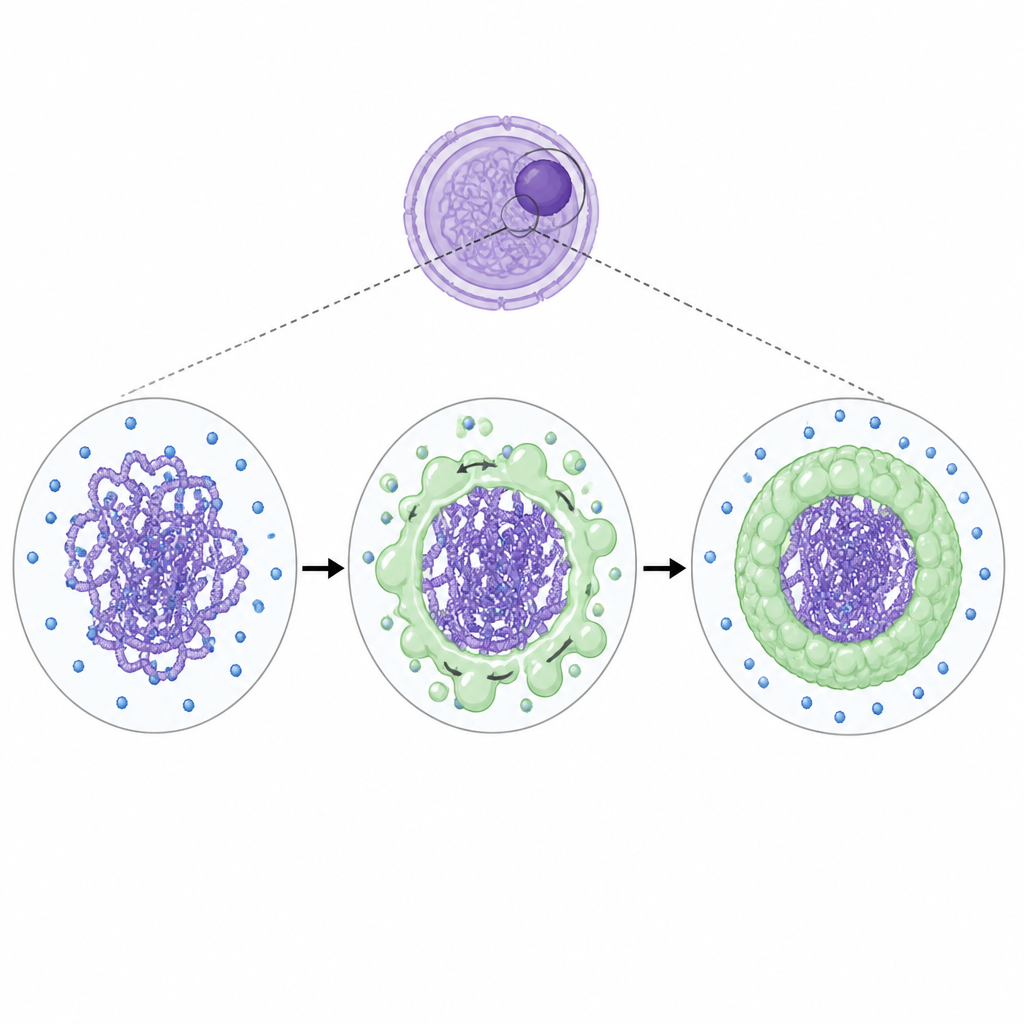
Неожиданное путешествие ядрышка Figure 1. Ядрышковые капли перемещаются на половые хромосомы, создавая временную зону тишины во время развития сперматозоидов.
Ядрышко наиболее известно как фабрика рибосом, где синтезируется и собирается рибосомная РНК. С помощью продвинутой 3D-микроскопии в тестах мышей исследователи обнаружили, что части ядрышка постепенно расщепляются и смещаются в сторону XY-тела во время пахитена. Два ядрышковых белка, NPM1 и SENP3, вместе с рибосомной РНК сначала формируют мелкие точки, а затем распространяются, покрывая XY-тело, после чего отступают на одну сторону от него. Тем временем обычное ядрышко в другой части ядра разрушается. Эта миграция наблюдалась и у мышей, и у людей и происходила только в мужских герминативных клетках, а не в женских, что указывает на полоспецифическую стратегию.
Ключевые ядрышковые участники поддерживают ход сперматогенеза
Чтобы проверить важность этих блуждающих фрагментов ядрышка, команда создала мышей, у которых NPM1 или SENP3 отсутствовали только в герминативных клетках. Внешне такие самцы выглядели нормально, но были полностью бесплодны. Их тесты были уменьшены, сперматоциты останавливались на стадии пахитена, а зрелые сперматозоиды практически отсутствовали. Детальное изображение хромосом показало, что в то время как аутосомы правильно сопрягались, X и Y часто имели деформированную форму или не складывались в обычную компактную структуру внутри XY-тела. В этих мутантных клетках подавленное XY-тело развивало аномальную полую, кольцевую форму, что указывает на то, что его внутренняя структура зависит от правильной работы NPM1 и SENP3.
Как ядрышковые капли отключают активность генов Figure 2. Жидкоподобные ядрышковые оболочки формируются вокруг области XY и вытесняют транскрипционный аппарат, подавляя их гены.
Исследователи также изучали активность генов напрямую. В нормальных клетках главный фермент, копирующий ДНК в РНК — РНК-полимераза II — практически отсутствует в XY-теле во время пахитена. В клетках, лишённых NPM1 или SENP3, этот фермент проникал в XY-тело, и многие гены на X и Y вновь неправильно активировались. Блокирование синтеза рибосомной РНК лекарствами вызывало схожие нарушения, что показывает, что сама РНК является частью механизма выключения. Биохимические тесты пояснили, как это работает: SENP3 модифицирует NPM1 так, что NPM1 начинает прочно связывать рибосомную РНК. Вместе они формируют жидкоподобные капли, которые ведут себя как отдельная фаза внутри ядра. В пробирочных экспериментах эти капли вытесняли компоненты транскрипционного аппарата к их внешним краям и снижали скорость синтеза РНК, что предполагает физический механизм, удерживающий полимеразу подальше от XY-хромосом.
Фазовое разделение как молекулярный выключатель
Затем команда изменила NPM1 так, чтобы он всё ещё связывался с РНК, но уже не мог образовывать капли. Мыши, несущие только эту версию NPM1, дефектную по фазовому разделению, снова оказались бесплодны: их сперматоциты останавливались, а гены, связанные с XY, реактивировались. Когда нормальный NPM1 возвращали в мутантные клетки, он собирался на XY-теле и восстанавливал вытеснение полимеразы, тогда как дефектный NPM1 этого сделать не мог. В совокупности результаты поддерживают модель, в которой датчики повреждений ДНК сначала помечают несопряжённые X и Y, затем привлекают NPM1, SENP3 и рибосомную РНК. Эти компоненты формируют жидкую оболочку вокруг XY-тела, которая физически вытесняет транскрипционный аппарат, выключая гены в нужный момент. Позднее более устойчивые химические метки на хроматине закрепляют это состояние тишины по мере созревания сперматозоидов. Для непрофессионального читателя главный вывод таков: крошечные подвижные капли внутри ядра помогают превратить половые хромосомы в защищённую зону тишины, и когда этот процесс нарушается, может развиться мужское бесплодие.
Цитирование: Li, M., Du, Z., Li, H. et al. Nucleolar migration regulates meiotic sex chromosome inactivation via phase separation during mammalian spermatogenesis.
Nat Commun17, 4485 (2026). https://doi.org/10.1038/s41467-026-70932-z