Clear Sky Science · ru
Генетическая архитектура аллостерического гормонального рецептора
Как крошечные изменения могут перестроить биологический переключатель
Наши клетки и клетки растений полагаются на молекулярные переключатели, которые обнаруживают химические сигналы и преобразуют их в действие. В этом исследовании изучается один такой переключатель — растительный гормональный рецептор PYL1 — и задаётся на первый взгляд простым вопросом: как изменения в его генетическом коде изменяют способ реакции на сигналы? Понимание этого важно не только для фундаментальной биологии, но и может помочь в проектировании культур, лучше переносящих засуху, или в создании рецепторов в виде пользовательских сенсоров.
Внимательный взгляд на сенсор стресса у растений
PYL1 помогает растениям реагировать на гормон стресса абсцизовую кислоту, важную для выживания в условиях сухости. В присутствии гормона PYL1 меняет конформацию и взаимодействует с другим белком, что запускает защитные ответы, включая активацию генов, связанных с реакцией на засуху. Как и многие рецепторы, PYL1 ведёт себя скорее как регулятор яркости, а не как простой вкл/выкл: по мере роста уровня гормона его активность описывается сигмовидной кривой — медленное включение, затем ускорение и в конце плато. Исследователи хотели выяснить, как каждая возможная замена одной буквы в последовательности PYL1 влияет на эту кривую: насколько чувствителен рецептор, каков его максимальный отклик и насколько резко он переключается между низкой и высокой активностью. 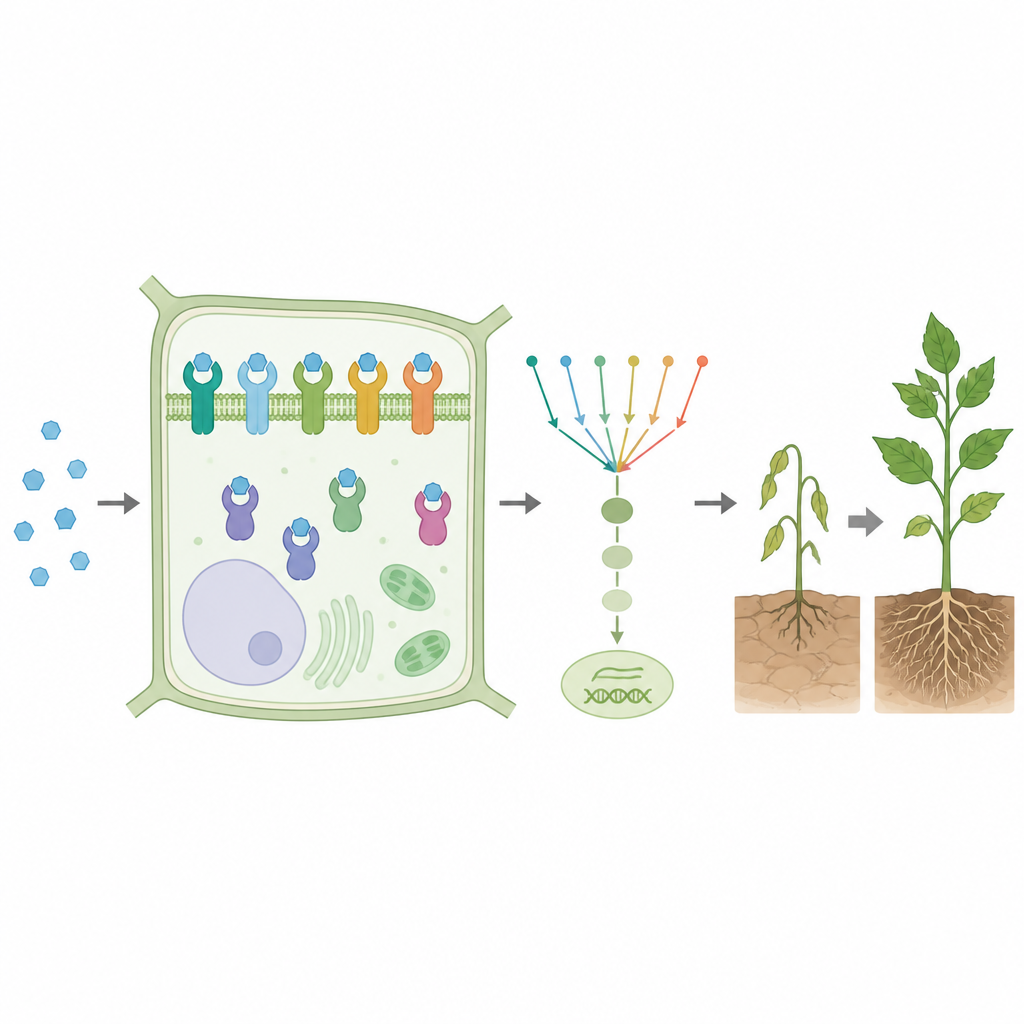
Одновременное измерение тысяч характеристик переключателя
Чтобы справиться с этой масштабной задачей, команда разработала высокопроизводительный метод, который они назвали GluePCA. Они сращивали PYL1 и его партнёрский белок с двумя половинками жизненно важного фермента в дрожжевых клетках. Когда PYL1 связывает партнёра в присутствии гормона, половинки фермента соединяются, фермент активируется, и дрожжи растут лучше. Вводя каждую возможную одиночную замену в PYL1 и подвергая дрожжи различным концентрациям гормона, исследователи могли с помощью секвенирования ДНК считывать, насколько хорошо работал каждый мутантный рецептор. Этот подход дал более 40 000 измерений и свыше 3 500 полных дозо‑ответных кривых, фактически создав карту того, как отдельные замены аминокислот настраивают поведение этого рецептора.
Как стабильность формирует силу сигнала
Данные показали, что почти 90 % миссенс‑мутаций, меняющих одну аминокислоту на другую, заметно изменяют кривую ответа PYL1. Многие мутации одновременно влияли на несколько характеристик: концентрацию гормона, необходимую для активации рецептора, базовую активность в отсутствие гормона и максимальную активность при высоких концентрациях гормона. Чтобы найти скрытую причину этих связанных изменений, команда независимо измерила, как каждая мутация влияет на стабильность и количество PYL1 с помощью отдельного анализа, оценивающего уровни рецептора через самосвязывание. Они обнаружили, что большинство мутаций делают рецептор менее стабильным, снижая его количество в клетках. Эти сдвиги стабильности объясняли почти три четверти разброса в сигнальных характеристиках: менее стабильные рецепторы, как правило, были менее чувствительны и имели более слабую максимальную активацию, тогда как более стабильные варианты демонстрировали более высокую базовую активность и плавное переключение.
Тонкая настройка и неожиданные новые типы переключателей
Однако стабильность была не единственным фактором. После математической корректировки на её влияние исследователи выделили группы положений в рецепторе, которые могли независимо настраивать отдельные параметры кривой ответа. Определённые области, удалённые от кармана для гормона, регулировали базовую активность; другие изменяли максимальный отклик; дополнительные участки вблизи полости связывания гормона ужесточали или ослабляли чувствительность. Такая модульная организация означает, что разные части структуры белка действуют как отдельные регуляторы формы его поведения. Удивительно, но небольшое число одиночных замен породило совершенно новые типы переключателей: некоторые мутанты инвертировали обычное поведение так, что гормон выключал взаимодействие вместо его включения, а другие создавали «полосовые‑стоп» паттерны, при которых сигнал отключался при промежуточных концентрациях гормона, но был активен при низких и высоких дозах. 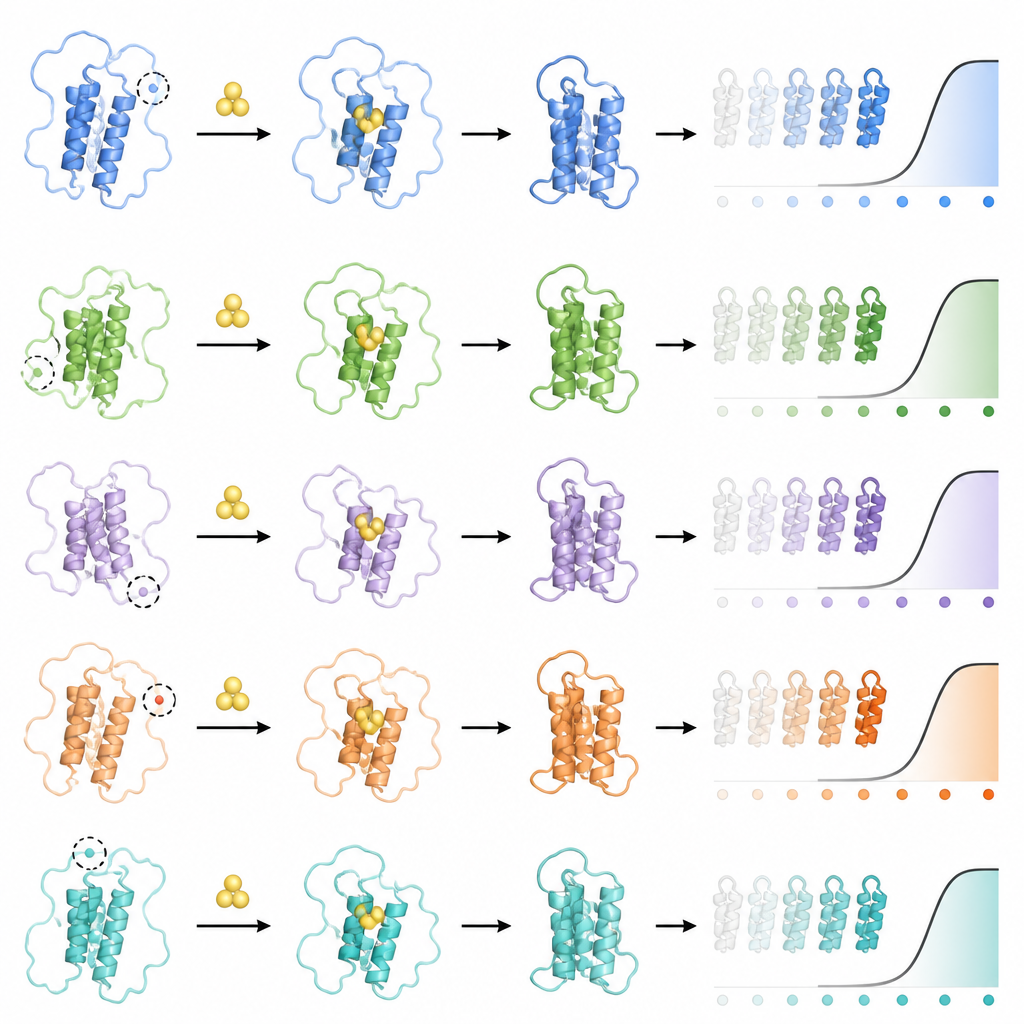
Почему это важно для эволюции и дизайна
Для неспециалиста ключевое сообщение заключается в том, что поведение рецептора гораздо более податливо, чем может показаться. Большинство одиночных замен в гене PYL1 тонко меняют способ, которым рецептор интерпретирует уровни гормона — в основном за счёт изменения стабильности белка, но также через целевые корректировки в отдельных структурных областях. Немногие редкие замены даже создают полностью новые типы переключателей. Это показывает, что у природы есть богатый инструментариум для эволюции новых сигнальных поведений и указывает на то, что учёные могли бы целенаправленно перепрограммировать такие рецепторы в качестве пользовательских сенсоров для сельского хозяйства, биотехнологии или медицины.
Цитирование: Stammnitz, M.R., Lehner, B. The genetic architecture of an allosteric hormone receptor. Nat Commun 17, 4735 (2026). https://doi.org/10.1038/s41467-026-70341-2
Ключевые слова: гормональный рецептор, аллостерия, стабильность белка, дозо‑ответ, сигнализация растения при засухе