Clear Sky Science · ru
Направленная динамика в энторинальной коре самцов мышей, вызванная поведенческими ограничениями
Как мозг понимает, куда он направлен
Ориентирование в пространстве зависит от внутреннего чувства направления, сформированного нейронами, которые активируются, когда животное смотрит в определённую сторону. В этом исследовании рассматривается на первый взгляд простой вопрос: являются ли эти «компасные» клетки жёстко запрограммированными или они могут менять свою роль, когда меняется способ нашего передвижения? Наблюдая за тысячами нейронов в ключевой области навигации у мышей, авторы показывают, что значительная часть этого направленного кодирования оказывается удивительно гибкой и формируется опытом.
Область мозга, которая картирует пространство
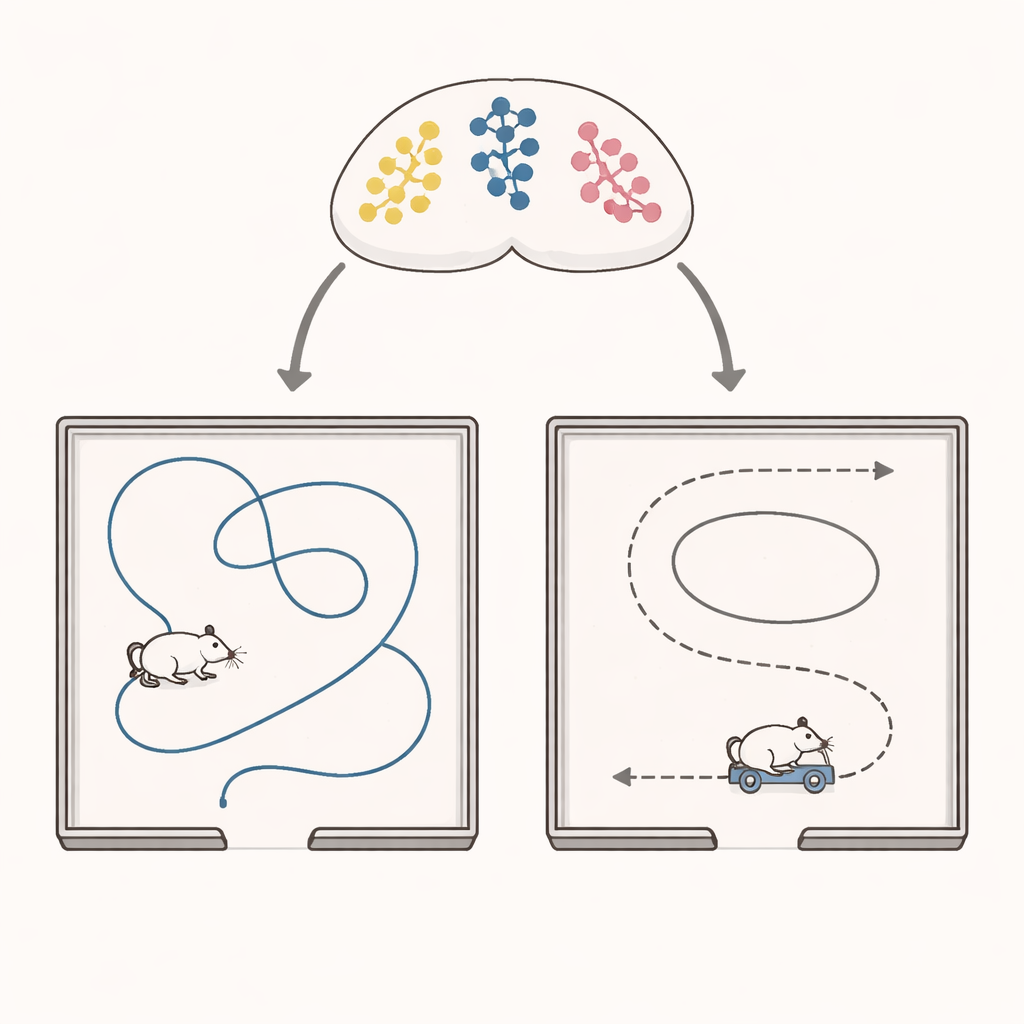
Глубоко в мозге медиальная энторинальная кора функционирует как часть навигационного узла. Она содержит несколько типов специализированных клеток, включая решётчатые клетки, кодирующие место, и клетки направления головы, реагирующие на ориентацию животного. До сих пор было неизвестно, фиксирована ли «должностная инструкция» клетки как клетки направления головы или её роль может переопределяться. Авторы применили двухфотонную кальциевую визуализацию, чтобы мониторить более 11 000 нейронов у мышей, которые исследовали квадратную арену. В одних сессиях животные свободно бродили; в других их головы были закреплены на небольшом автономном тележке, который перемещал их по тому же пространству, меняя лишь способ передвижения, но не маршруты.
Когда свободное блуждание превращается в управляемую поездку
Учёные сначала сравнили направленные сигналы при свободном движении и при «помощной навигации» на тележке в арене с множеством ориентиров — сильными визуальными и обонятельными метками. Удивительно, но общее направленное тонкое кодирование в энторинальной коре стало более острым, информативным и стабильным, когда мыши находились на тележке. Но это улучшение скрывало поразительную реорганизацию. Одна группа нейронов, чья активность явно соответствовала направлению головы во время свободного исследования, потеряла это кодирование на тележке. Вторая группа, ранее непримечательная, приобрела сильную направленную селективность только во время помощной навигации. Небольшая третья группа сохраняла надёжную селективность в обоих условиях. Анализы декодирования подтвердили, что популяционный сигнал о направлении работал лучше всего, когда декодер обучался и тестировался в одном и том же режиме навигации, что указывает на реальную переконфигурацию набора активных клеток.
Ограничения, ориентиры и новая карта
Чтобы выяснить, что вызывает этот переход, команда изменила окружение и движение тележки. В «бедной ориентирами» арене, лишённой большей части визуальной структуры, помощная навигация больше не улучшала направленное кодирование: меньше клеток приобретали селективность, и карты становились менее стабильными. Изменение профиля скорости тележки, однако, имело мало влияния; новые клетки направления головы сохраняли схожие предпочитаемые направления даже при медленном или быстром движении тележки. Это указывает на то, что ключевым фактором для привлечения новых направленных клеток при ограничениях являются богатые внешние сенсорные ориентиры, а не простые статистики движения. В то же время подкласс инвариантных клеток сохранял координированную активацию во всех условиях, что предполагает, что они образуют более жёстко заданный, аттрактороподобный каркас направления.
Обучение второму компасу
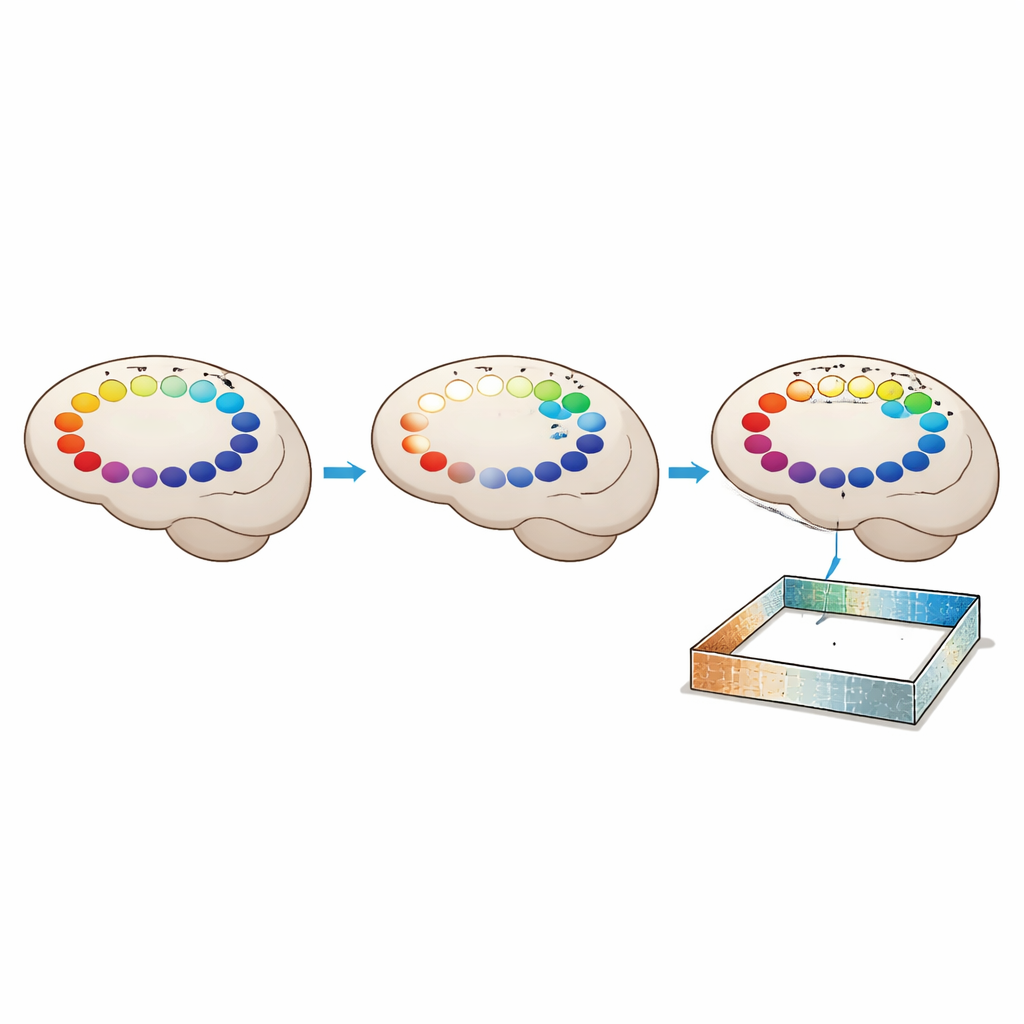
Авторы затем выяснили, как быстро формируется этот код для помощной навигации. У мышей, впервые испытавших тележку, качество направленных карт и способность декодировать направление по ним улучшались постепенно в течение одной сессии, длящейся всего несколько минут. В последующих «тренированных» сессиях эти показатели уже были высоки и мало менялись, что свидетельствует о том, что новый код был выучен и сохранён. Во время помощной навигации многие клетки направления головы также начали нести информацию о том, где именно в арене находится животное, а не только о том, куда оно обращено. Их пространственные поля активности склонялись к скоплению у стен, и их предпочитаемые направления чаще указывали на ближайшую стену в условиях с множеством ориентиров, чем в арене с их недостатком. Это указывает на то, что при поведенческих ограничениях энторинальная карта связывает направление и место через близкие сенсорные ориентиры.
Гибкий внутренний компас
Для неспециалиста основной вывод заключается в том, что внутренний компас мозга не представляет собой один фиксированный механизм. Скорее он сочетает небольшой стабильный ядро клеток направления с более крупным адаптивным пулом, чьи роли могут переключаться в зависимости от того, как животное движется и что оно ощущает. Когда движение головы ограничено, но доступны богатые ориентиры, привлекаются новые нейроны, помогающие поддерживать надёжное чувство направления, и они быстро учатся связывать это чувство с конкретными местами и стенами. Эта работа предполагает, что наша навигационная система может хранить несколько карт одного и того же места, выбирая ту, которая лучше соответствует текущему способу передвижения по миру.
Цитирование: Liu, R., Hao, J., Zhang, X. et al. Directional dynamics in the entorhinal cortex of male mice driven by behavioral constraints. Nat Commun 17, 3679 (2026). https://doi.org/10.1038/s41467-026-70289-3
Ключевые слова: нейроны направления головы, энторинальная кора, пространственная навигация, сенсорные сигналы, нейронная пластичность