Clear Sky Science · pl
Interakcja między dawką DYRK1A a aneuploidią wywołaną neuropatologią w zespole Downa
Dlaczego to badanie ma znaczenie dla rodzin i opiekunów
Osoby z zespołem Downa często mają trudności w nauce i zwiększone ryzyko wczesnej utraty pamięci, lecz przyczyny wysokiej wrażliwości komórek mózgowych pozostawały niejasne. To badanie wykorzystuje modele z ludzkich komórek macierzystych, aby odsłonić, jak dwa ukryte problemy wewnątrz komórek mózgu współdziałają, uszkadzając neurony, i sugeruje połączoną strategię terapeutyczną, która mogłaby w przyszłości pomóc chronić funkcje poznawcze i pamięć osób z zespołem Downa. 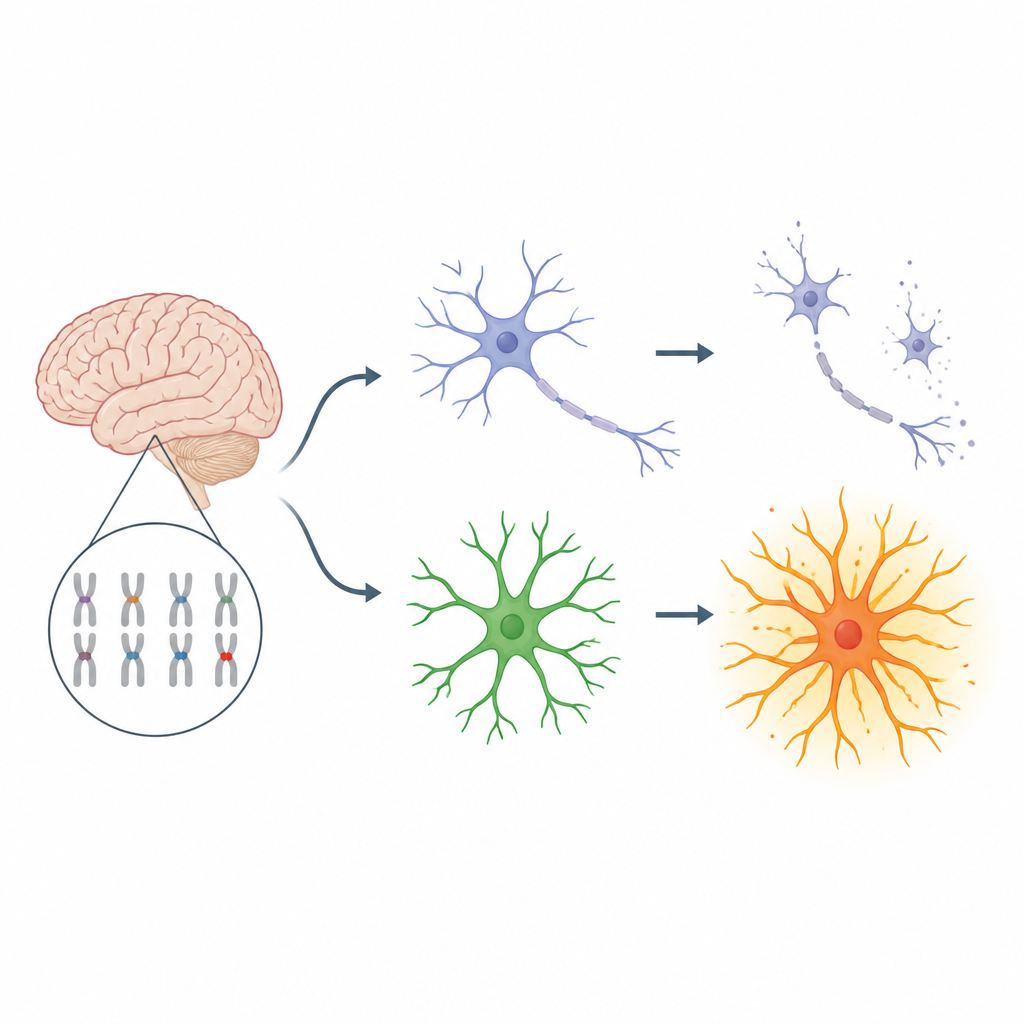
Dwa rodzaje komórek mózgowych uwikłane w szkodliwe partnerstwo
Mózg opiera się na ścisłej współpracy między neuronami, które przekazują sygnały, a astrocytami — gwiaździstymi komórkami wspierającymi, które kierują i chronią neurony. W zespole Downa dodatkowa kopia chromosomu 21 zaburza tę równowagę: jest mniej neuronów i w przybliżeniu dwa razy więcej astrocytów niż zwykle, a astrocyty mają tendencję do utrzymywania przewlekle „włączonego” stanu. Badacze postawili pytanie, jak stres wewnątrz komórek trisomicznych i dodatkowe kopie niektórych genów z chromosomu 21 mogą się łączyć w obrębie tego partnerstwa neuron–astrocyt, powodując długotrwałe uszkodzenia.
Zestresowane neurony obciążone zbrylonymi białkami
Wykorzystując indukowane komórki macierzyste pluripotentne od osób z trisomią 21, 18 i 13, zespół hodował neurony w laboratorium i porównywał je z dopasowanymi komórkami skorygowanymi do zwykłej liczby chromosomów. We wszystkich trzech trisomiach neurony wykazywały silne oznaki stresu wewnętrznego: białka źle się fałdowały i tworzyły agregaty, a więcej komórek przechodziło zaprogramowaną śmierć komórkową. Kluczowe białko powiązane z chorobą Alzheimera, tau, występowało w nadmiernie fosforylowanej, skłonnej do agregacji formie, która przenikała do płynu otaczającego neurony. Leczenie neuronów chemicznym „chaperonem” 4‑fenylobutyratem zmniejszyło te grudki białkowe i obniżyło ilość uwalnianego zestresowanego tau, pokazując, że sama nierównowaga chromosomalna może wywołać wspólny rodzaj toksycznego nagromadzenia białek. 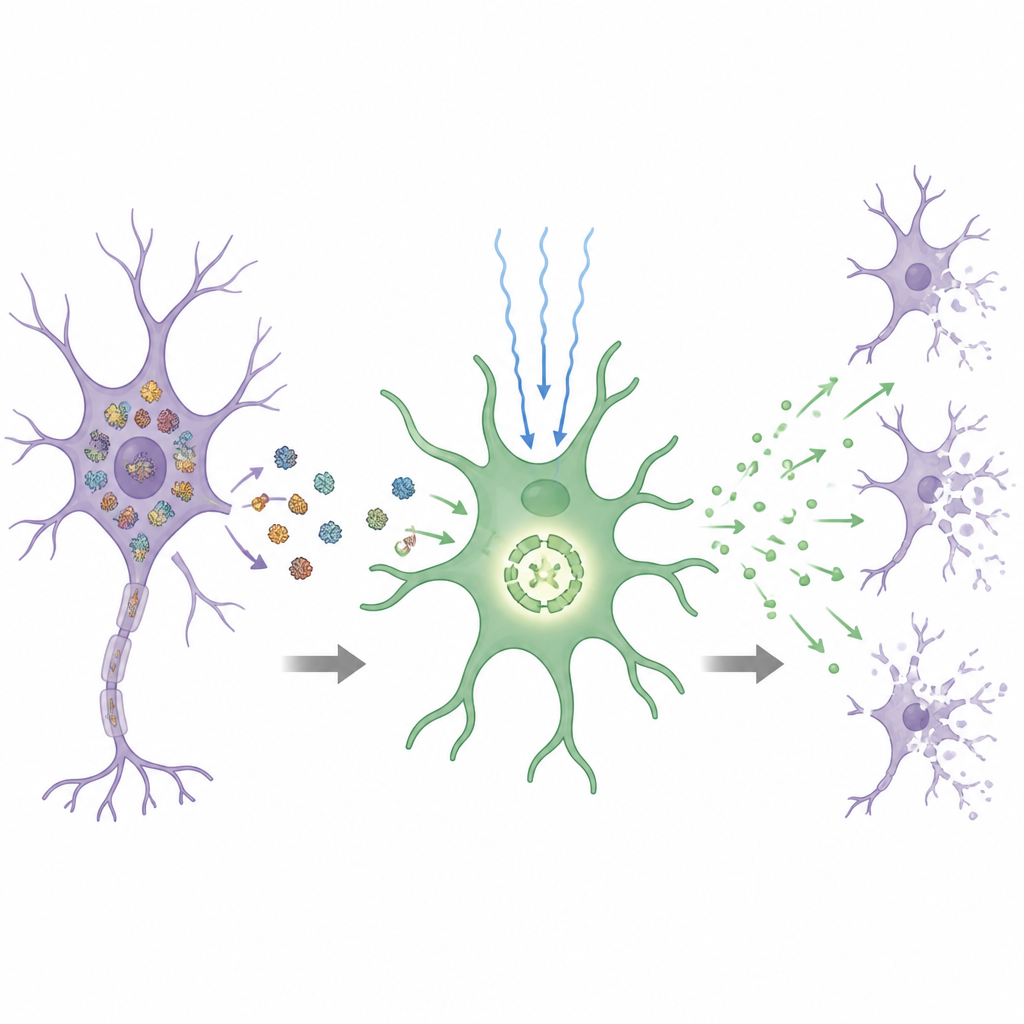
Astrocyty w zespole Downa stają się naturalnymi rozpalaczami
Historia wyglądała inaczej w przypadku astrocytów. Tylko astrocyty z trisomią 21 wykazywały wyjątkowo szybki wzrost i silny profil zapalny, wskazując na specyficzny efekt dodatkowego chromosomu 21, a nie ogólny stres chromosomalny. Te astrocyty miały wewnętrznie aktywowany czujnik niebezpieczeństwa zwany inflammasomem NLRP3, który normalnie pomaga kontrolować uwalnianie substancji zapalnych. Nawet bez standardowych sygnałów inicjujących, astrocyty z trisomią 21 produkowały wysokie poziomy cząsteczek zapalnych, takich jak interleukina‑1β, a tę odpowiedź można było stłumić lekami blokującymi NLRP3. Gdy neurony hodowano razem z astrocytami z trisomią 21, agregacja białek i śmierć neuronów wzrosły gwałtownie, co pokazuje, że te reaktywne komórki wspierające mogą zmienić się z opiekunów w sprawców uszkodzeń.
Vicious loop napędzany jednym dodatkowym genem
Badacze następnie pytali, który gen z chromosomu 21 popycha astrocyty w kierunku tego stanu zapalnego. Studiując linię komórkową pozbawioną jedynie małego krytycznego regionu chromosomu 21, a następnie selektywnie wyciszając pojedyncze geny, wytypowali DYRK1A — gen już znany z roli w rozwoju mózgu. Dodatkowa kopia DYRK1A zwiększała kluczowe szlaki sygnałowe w astrocytach prowadzące do NLRP3, podczas gdy zmniejszenie aktywności DYRK1A przez korekcję genetyczną lub leki wyciszało ekspresję NLRP3 i jego produktów zapalnych. Jednocześnie medium pobrane od zestresowanych neuronów trisomicznych dodatkowo potęgowało odpowiedź zapalną astrocytów z trisomią 21, a efekt ten zanikał, gdy stres białkowy neuronów został złagodzony przez 4‑fenylobutyrat. Razem te odkrycia ujawniają pętlę napędzającą postęp: stres chromosomalny powoduje, że neurony uwalniają toksyczne formy białek, które aktywują astrocyty uwrażliwione przez DYRK1A, a te z kolei wydzielają sygnały zwiększające skłonność neuronów do obumierania.
Połączenie dwóch dźwigni dla lepszej ochrony neuronów
Ponieważ pętla uszkodzeń ma dwa główne napędy, zespół przetestował dwutorową strategię. Skorygowanie liczby kopii DYRK1A jedynie w astrocytach zmniejszyło sygnalizację zapalną i obniżyło stres neuronalny, ale neurony z trisomią 21 nadal częściej umierały niż skorygowane kontrolne. Gdy badacze połączyli korekcję DYRK1A w astrocytach z leczeniem 4‑fenylobutyratem łagodzącym stres białkowy w neuronach, zarówno agregacja białek, jak i śmierć neuronów wróciły blisko poziomów prawidłowych. Dla odbiorcy nieprofesjonalnego wniosek jest taki, że w zespole Downa utrata neuronów może wynikać ze współdziałania zestresowanych neuronów i nadaktywnych komórek wspierających, i że uspokojenie obu stron jednocześnie — poprzez wyciszenie DYRK1A w astrocytach oraz złagodzenie stresu białkowego w neuronach — mogłoby stać się podstawą przyszłych terapii mających na celu ochronę zdrowia mózgu.
Cytowanie: Nambara, T., Lee, J.Y., Minami, M. et al. Interplay between DYRK1A dosage and aneuploidy-induced neuropathology in Down syndrome. Commun Biol 9, 660 (2026). https://doi.org/10.1038/s42003-026-09902-6
Słowa kluczowe: Zespół Downa, neurony, astrocyty, zapalenie mózgu, agregaty białkowe