Clear Sky Science · pl
Równoległe ekrany CRISPR ujawniają szlaki kontrolujące poziomy receptora atraktonu FPR1 na powierzchni komórek
Jak komórki odpornościowe dostrajają swoją czułość
Nasz układ odpornościowy polega na wysuniętych na pierwszą linię komórkach zwanych neutrofilami, które pędzą do miejsc zakażenia, kierowane chemicznymi śladami. Aby uniknąć zarówno ospałej obrony, jak i wymykającego się spod kontroli zapalenia, komórki te muszą ostrożnie kontrolować, ile „receptorów węchowych” znajduje się na ich powierzchni. W tym badaniu zadano proste, lecz istotne pytanie: jak neutrofile decydują, kiedy ściągnąć te receptory do wnętrza komórki, a kiedy wysłać więcej na zewnątrz, i co dzieje się w komórce podczas tego ciągłego ruchu?
Uważny strażnik na neutrofilach
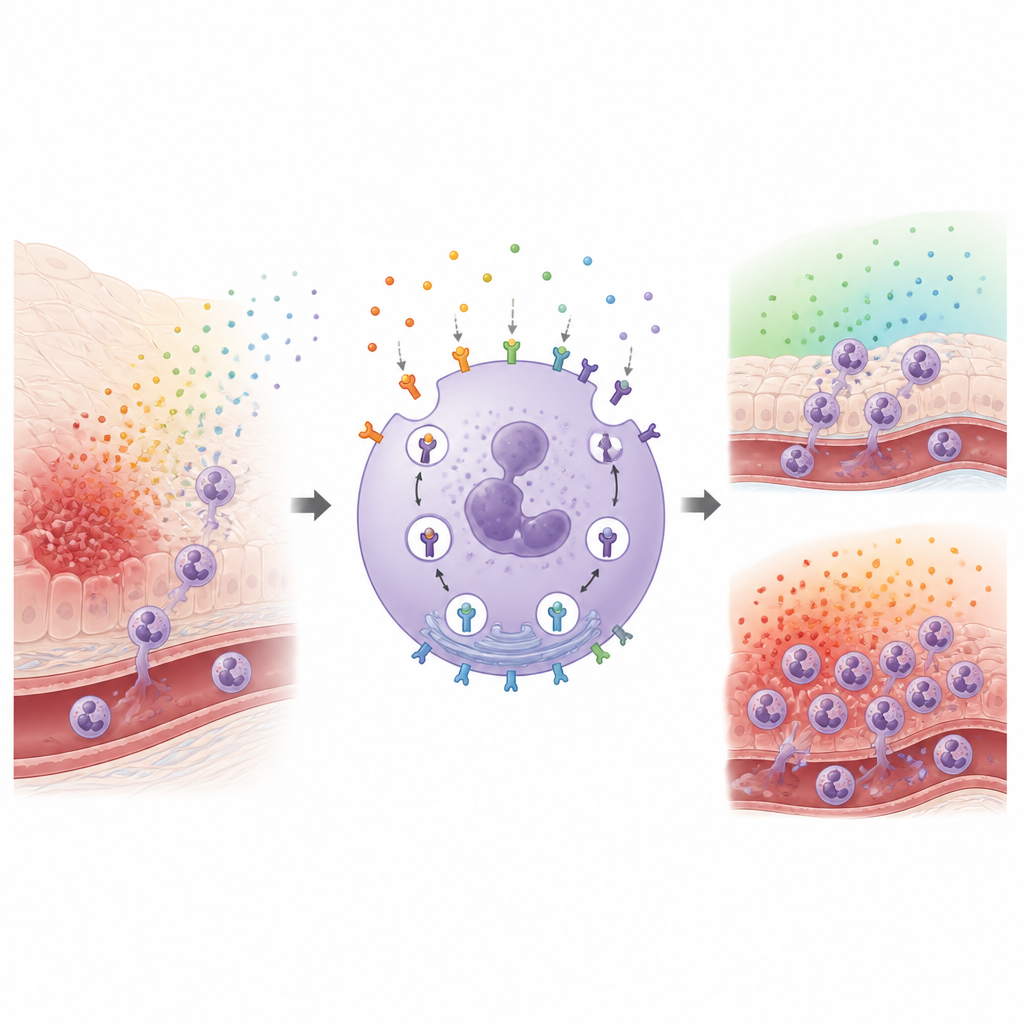
Kluczowym graczem w tej historii jest receptor zwany FPR1, który znajduje się na powierzchni neutrofili i rozpoznaje drobne fragmenty wydzielane przez bakterie oraz uszkodzone tkanki. Gdy FPR1 odbiera te sygnały alarmowe, pomaga neutrofilom migrować w stronę zagrożenia i uruchamiać mechanizmy zdolne zabijać drobnoustroje, a jednocześnie uszkadzać zdrowe tkanki. Liczba receptorów FPR1 na powierzchni silnie wpływa na czułość neutrofila. Po aktywacji wiele receptorów jest ściąganych do wnętrza, obniżając czułość, podczas gdy w innych warunkach więcej receptorów trafia na powierzchnię, przygotowując komórkę do reakcji. Szczegółowe mechanizmy dodawania i usuwania FPR1 z powierzchni komórki pozostawały jednak zaskakująco niejasne.
Pomiary ruchu receptorów w żywych komórkach
Naukowcy najpierw udoskonalili sposób obserwowania przemieszczania się FPR1 w dużych populacjach komórek jednocześnie. Użyli linii komórkowej przypominającej neutrofile i znakowali powierzchniowy FPR1 fluorescencyjnymi przeciwciałami, a następnie stymulowali receptory bakteryjnym mimetykiem. Śledząc, jak szybko sygnał powierzchniowy maleje, mogli wnioskować, jak szybko receptory są internalizowane. Potwierdzili te pomiary mikroskopią, która pokazała akumulację fluorescencyjnego atraktonu wewnątrz komórek w miarę zanikania barwienia powierzchni. W ciągu kilku minut większość FPR1 zniknęła z błony, co ujawniło, że ten receptor jest usuwany szybko i efektywnie, a jego część jest później recyrkulowana z powrotem na powierzchnię.
Odkrywanie równoległych dróg do wnętrza komórki
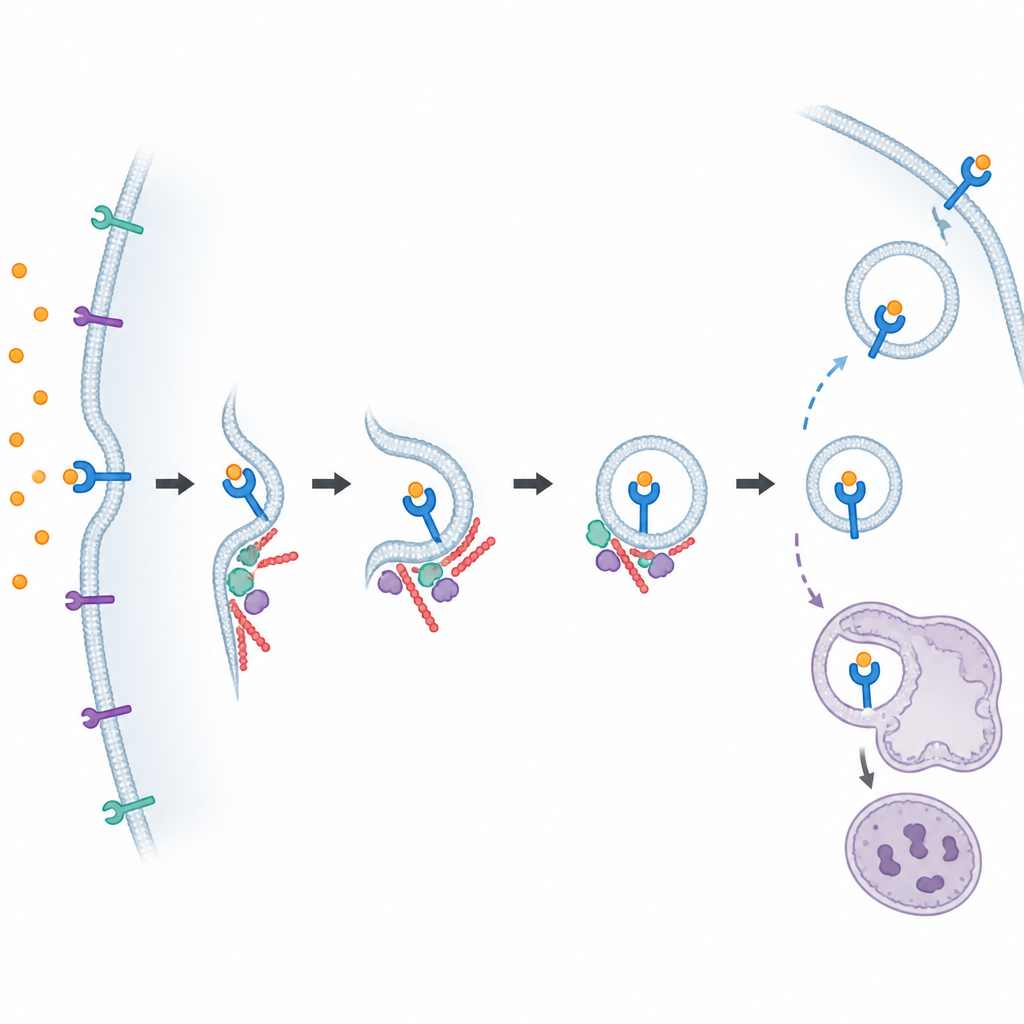
Następnie zespół przyjrzał się znanym białkom regulacyjnym. Pokażali, że kilka enzymów celujących w receptory, zwanych GRK, działa wspólnie, znakując FPR1 tak, by mógł zostać ściągnięty do wnętrza, a że dwa białka adapterowe znane jako beta-arrestyny wspomagają ten proces. Jednak nawet gdy obie beta-arrestyny zostały usunięte, FPR1 nadal częściowo się internalizował, co sugeruje, że przynajmniej jedna dodatkowa droga do wnętrza działa równolegle. Aby systematycznie przeszukać wszystkich uczestników, badacze sięgnęli po ognogenomowe ekrany CRISPR, zakłócając niemal każdy gen w genomie w ogromnych pulach komórek. Przeprowadzili dwa powiązane ekrany: jeden wychwytywał geny wpływające na poziomy FPR1 w komórkach spoczynkowych, a drugi wychwytywał geny zmieniające poziomy FPR1 po stymulacji, co pozwoliło im rozróżnić problemy w wytwarzaniu, transporcie, usuwaniu lub recyklingu receptora.
Ukryte mechanizmy recyklingu receptora
Porównując te ekrany, autorzy zmapowali sieć szlaków kontrolujących ruch FPR1. Wyróżnili duże zespoły białkowe, które pomagają sfałdować FPR1, przenieść go przez wewnętrzne szlaki komórkowe i sortować w endosomach, które są stacjami tranzytowymi dla napływającego ładunku. Niektóre kompleksy, takie jak retromer, retriever i CCC, wydawały się wpływać zarówno na podstawową ilość FPR1 na powierzchni, jak i na to, co dzieje się z nim po internalizacji. Badanie wskazało także na mechanizmy przesuwające ziarna magazynujące do błony, umożliwiając nagłe dostawy FPR1, gdy komórki napotkają atrakton. Ta zintegrowana perspektywa pokazuje, że poziomy receptorów nie są kontrolowane jednym przełącznikiem, lecz wielowarstwowym systemem produkcji, trasowania i recyklingu.
Nowe molekuły kierujące pobieraniem receptorów
Wśród licznych trafień szczególnie ważne okazały się dwa wcześniej niedoceniane białka, istotne dla ściągania FPR1 z powierzchni. Jedno, mDia1, pomaga budować prostolinijne filamenty aktynowe, część wewnętrznego rusztowania komórki. Drugie, ARF6, to mały przełącznik molekularny wpływający na to, jak błona się wygina i jak powstają pęcherzyki. Gdy zespół chemicznie zahamował mDia1 lub ARF6, albo genetycznie usunął ARF6, komórki przypominające neutrofile oraz pierwotne ludzkie neutrofile nie były w stanie prawidłowo internalizować FPR1. Dalsze eksperymenty sugerowały, że ARF6 uczestniczy szczególnie w odnodze szlaku niezależnej od beta-arrestyn, wzmacniając koncepcję, że FPR1 korzysta z więcej niż jednej drogi internalizacji.
Dlaczego ta mapa ruchu komórkowego ma znaczenie
Dla odbiorców niebędących specjalistami płynie stąd prosty wniosek: neutrofile regulują swój „pokrętło głośności” dla sygnałów zagrożenia, ściśle zarządzając liczbą receptorów FPR1 wystawionych na powierzchni, korzystając z kilku nakładających się ścieżek. Badanie dostarcza mapy białek i kompleksów przenoszących FPR1 na błonę i z niej oraz ujawnia, że czynniki budujące aktynę, takie jak mDia1, oraz regulator błony ARF6 są kluczowymi elementami tego systemu. Zrozumienie tych dróg może w przyszłości pomóc naukowcom zaprojektować terapie łagodzące szkodliwe zapalenie lub wykorzystać szybki internalizację receptorów do ukierunkowanego dostarczania leków, poprzez delikatne manipulowanie wewnętrzną kontrolą ruchu komórek zamiast całkowitego wyłączania sygnałów.
Cytowanie: Akdoğan, E., Lundgren, S.M., Kamber, R.A. et al. Parallel CRISPR screens reveal pathways controlling the cell surface levels of the attractant receptor FPR1. Commun Biol 9, 668 (2026). https://doi.org/10.1038/s42003-026-09878-3
Słowa kluczowe: neutrofile, receptor FPR1, ekran CRISPR, endocytoza receptora, sygnalizacja immunologiczna