Clear Sky Science · pl
Fosforylacja β‑tubuliny przez Chk1 jest wymagana do prawidłowego tworzenia wrzeciona podczas podziału komórkowego
Jak komórki utrzymują porządek w swoim materiale genetycznym
Za każdym razem, gdy komórka się dzieli, stoi przed wysokostawkowym wyzwaniem inżynieryjnym: musi zbudować maleńką maszynę, która dokładnie rozdzieli skopiowane chromosomy, tak by każda komórka potomna otrzymała właściwy ładunek genetyczny. Gdy ta maszyna, zwana wrzecionem, zawodzi, skutki mogą obejmować wady rozwojowe, zaburzenia rozwoju lub raka. W tym badaniu odkryto nieoczekiwany sposób, w jaki dobrze znany białkowy strażnik uszkodzeń DNA, Chk1, pełni drugorzędną rolę kierownika budowy samego wrzeciona.
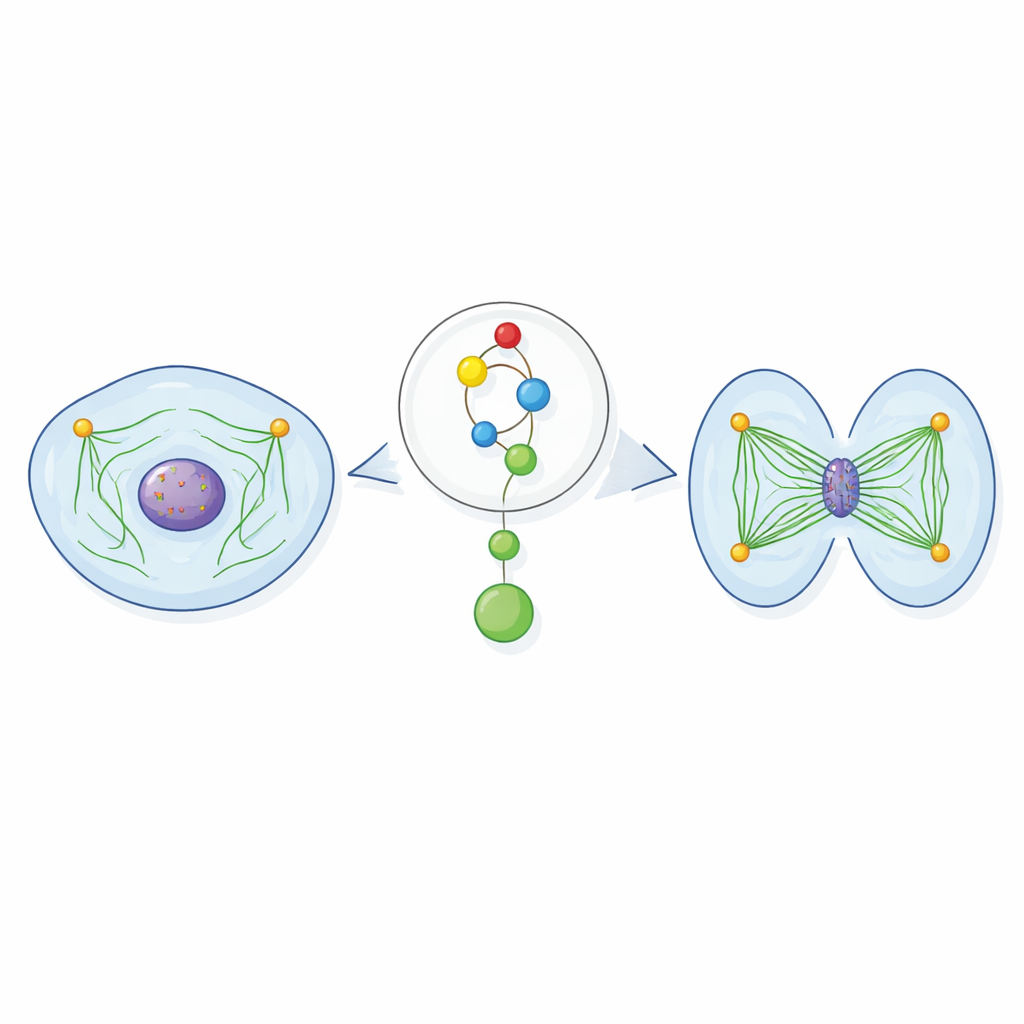
System lin i bloczków komórki
Podczas podziału komórkowego długie włókna białkowe zwane mikrotubulami tworzą wrzeciono — bipolarną strukturę, która chwyta chromosomy i przeciąga je na przeciwległe strony komórki. Te włókna wyrastają z centrów organizujących zwanych centrosomami i przyczepiają się do określonych miejsc na chromosomach. Aby to działało, wrzeciono potrzebuje wystarczającej liczby mikrotubul ułożonych w odpowiedniej geometrii, by uchwycić i przemieścić każdy chromosom. Jeśli wrzeciono jest zbyt rzadkie lub niestabilne, chromosomy mogą zostawać w tyle, źle się przyłączać lub trafiać do niewłaściwej komórki potomnej — stan znany jako aneuploidia, silnie związany z rakiem.
Strażnik DNA z dodatkowym zadaniem
Chk1 jest najbardziej znany jako element systemu odpowiedzi na uszkodzenia DNA: gdy DNA jest uszkodzone lub replikacja zatrzymuje się, Chk1 wstrzymuje cykl komórkowy, by umożliwić naprawę. Autorzy tego artykułu zastanawiali się, czy Chk1 może także działać podczas normalnego, niezakłóconego podziału komórki. Przez obniżenie poziomów Chk1 lub zablokowanie jego aktywności w kilku liniach komórek kręgowców zauważyli, że wrzeciona nadal się formują, ale są wyraźnie cieńsze, z mniejszą liczbą mikrotubul promieniujących z centrosomów. Te defekty pojawiały się nawet przy nienaruszonym DNA i przy utrzymaniu stałych innych znanych kroków kontrolowanych przez Chk1, takich jak wejście w mitozę czy aktywacja innego regulatora mitotycznego, Aurory B. To wykazało, że Chk1 ma odrębną, bezpośrednią rolę w budowie solidnego wrzeciona.
Włączanie tubuliny, by zbudować silniejsze wrzeciono
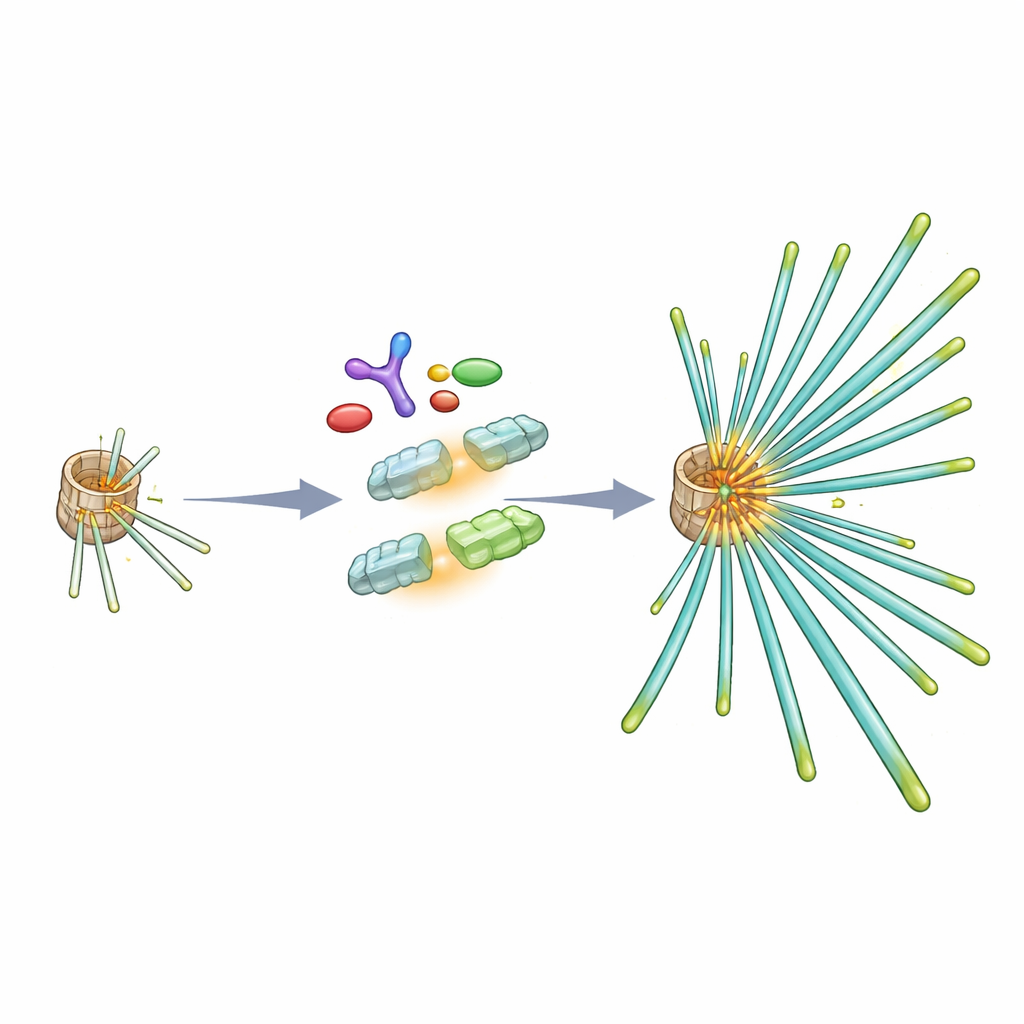
Aby zrozumieć, jak Chk1 wzmacnia wrzeciono, badacze zwrócili się ku tubulinie, podstawowemu budulcowi mikrotubul. Odkryli, że Chk1 fizycznie kojarzy się z tubuliną i może chemicznie modyfikować podjednostkę β‑tubuliny w konkretnym aminokwasie (treonina 285) w eksperymentach in vitro. W dzielących się komórkach odpowiadająca modyfikacja pojawia się wokół centrosomów specyficznie we wczesnej i środkowej fazie mitozy, właśnie wtedy gdy mikrotubule są nukleowane. Gdy komórki zaprojektowano tak, by produkowały wersję β‑tubuliny, której nie można zmodyfikować w tym miejscu, ich wrzeciona przypominały stan pozbawiony Chk1: mikrotubule były mniej gęste, odrastały wolniej po rozmontowaniu przez zimno i mniej stabilnie przyłączały się do chromosomów. Wersja „fosfomimetyczna” β‑tubuliny, zachowująca się tak, jakby była trwale zmodyfikowana, mogła natomiast naprawić defekty wrzeciona spowodowane zablokowaniem Chk1 lub jego aktywatora uprzedniego, ATR.
Utrzymanie czasu i symetrii podziału
Wrzeciona pozbawione prawidłowo zmodyfikowanej β‑tubuliny robią coś więcej niż tylko wyglądają na słabe — źle się zachowują. Chromosomy w tych komórkach często nie ustawiają się równo na linii środkowej komórki i wykazują zwiększoną nierówną segregację podczas podziału. Wbudowany mechanizm bezpieczeństwa monitorujący przyłączenia wrzeciona pozostaje aktywny dłużej, opóźniając początek anafazy. Obrazowanie komórek na żywo ujawniło, że komórki z niemodyfikowalną β‑tubuliną potrzebują więcej czasu, by przejść od zaokrąglenia się do faktycznego rozdzielenia chromosomów. Nieprawidłowe działanie wrzeciona wpływa też na ostateczne rozcięcie: gdy Chk1 lub modyfikacja β‑tubuliny są zaburzone, wrzeciona mają tendencję do ustawiania się poza osią, powodując, że komórka zaciska się w nieodpowiednim miejscu i powstają komórki potomne o nierównej wielkości, co może zaburzać równowagę zawartości komórkowej i sygnalizacji.
Przełączenie ścieżki odpowiedzi na uszkodzenia w celu budowy wrzeciona
Uprzednio wobec Chk1, badanie pokazuje, że trzy białka zwykle kojarzone z uszkodzeniami DNA — ATR, jego partner ATRIP oraz rusztowanie TopBP1 — gromadzą się w centrosomach podczas mitozy. ATRIP jest kluczowy dla sprowadzenia zarówno ATR, jak i TopBP1 do tych struktur. Jeżeli ich interakcje zostaną zakłócone, Chk1 nie jest już właściwie aktywowany w centrosomach, β‑tubulina nie zostaje zmodyfikowana w krytycznym miejscu, a mikrotubule wrzeciona ponownie są rzadkie. Zmodyfikowana β‑tubulina preferencyjnie trafia do frakcji polimeryzowanych mikrotubul, co sugeruje, że ta chemiczna modyfikacja pomaga podjednostkom tubuliny efektywnie włączać się do rosnących włókien i wspiera gęstą sieć potrzebną do pewnego przemieszczania chromosomów.
Dlaczego to ma znaczenie dla zdrowia i raka
Autorzy konkludują, że komórki przekształcają modul sygnalizacji uszkodzeń DNA w centrosomach, by dopracować konstrukcję wrzeciona podczas normalnego podziału. Poprzez modyfikację pojedynczego, ewolucyjnie zachowanego miejsca w β‑tubulinie, Chk1 sprzyja wydajnej nukleacji mikrotubul, terminowemu przebiegowi mitozy, dokładnej segregacji chromosomów, równemu rozmiarowi komórek potomnych i intensywnemu rozmnażaniu komórek. Ponieważ błędy w tych procesach są cechami charakterystycznymi chorób genetycznych i ewolucji nowotworowej, zrozumienie tej ścieżki budowy wrzeciona może otworzyć nowe możliwości terapii przeciwnowotworowej, na przykład przez łączenie inhibitorów ATR lub Chk1 z istniejącymi lekami celującymi w mikrotubule.
Cytowanie: Boutakoglou, N., Petsalaki, E., Balafouti, S. et al. β-tubulin phosphorylation by Chk1 is required for normal spindle formation during cell division. Commun Biol 9, 608 (2026). https://doi.org/10.1038/s42003-026-09862-x
Słowa kluczowe: wrzeciono mitotyczne, Chk1, beta tubulina, segregacja chromosomów, sygnalizacja ATR