Clear Sky Science · pl
Sieci skrzyżowanego filamentarnego F-aktyny regulują zależną od obciążenia konwersję energii
Jak komórki zamieniają paliwo w siłę
Każdy ruch twojego ciała, od bicia serca po zmianę kształtu rozwijającego się embrionu, zależy od tego, że komórki zamieniają chemiczne paliwo w siłę mechaniczną. W artykule tym przyglądamy się temu procesowi na bardzo drobnym poziomie, zadając pozornie proste pytanie: jak wewnętrzne „rusztowanie” komórki kontroluje, jak wydajnie przekształca energię z ATP — paliwa komórkowego — w ruch i siłę? Autorzy pokazują, że małe białka łączące filamenty aktynowe działają jak ukryte regulatory, strojąc, jak mocno ciągną nanomotory i ile energii przy tym zużywają.
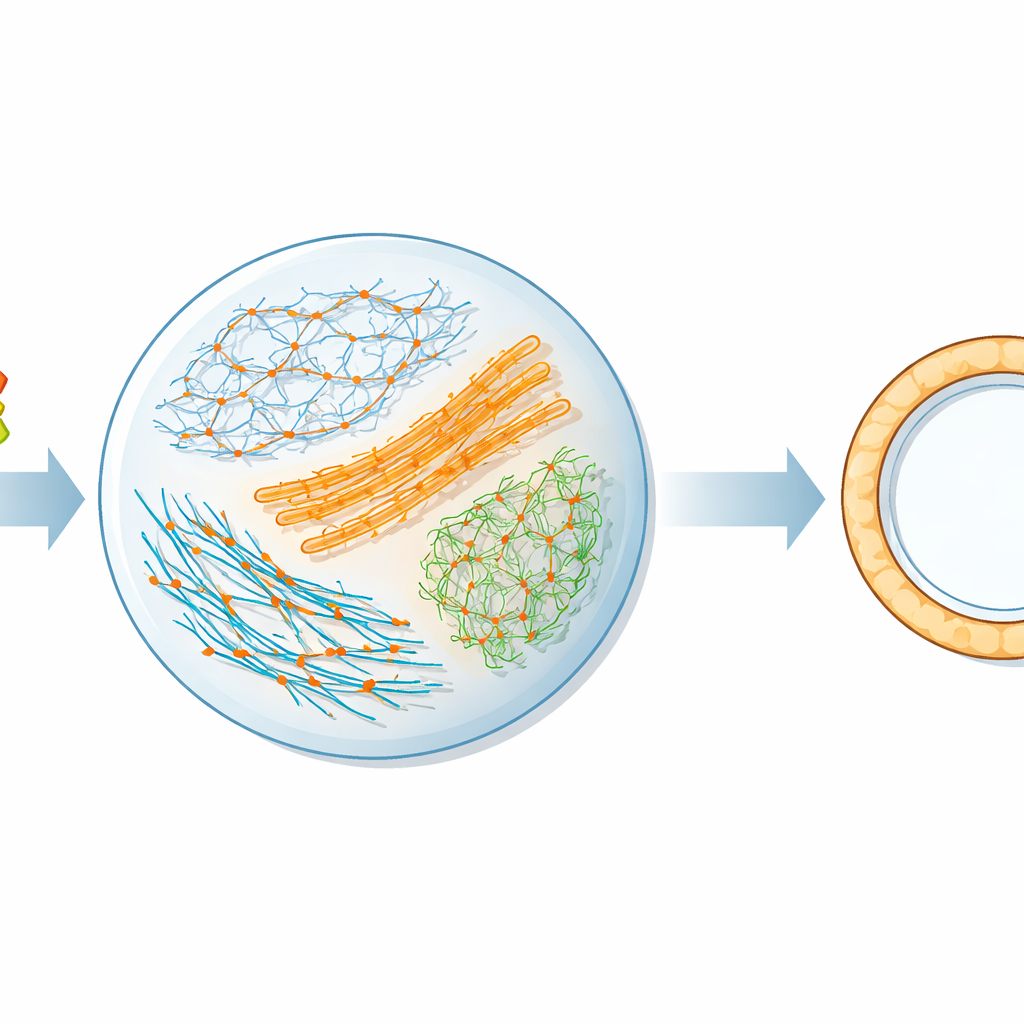
Budując małe laboratorium w kropli
Aby zbadać ten proces w kontrolowanych warunkach, badacze skonstruowali miniaturową wersję maszyny generującej siłę w komórce. Wymieszali oczyszczone filamenty aktyny, które tworzą wewnętrzny szkielet komórki, z motorami miozynowymi, które pełzają po aktynie i generują siły skurczowe. Mieszankę zamknięto w mikroskopijnych kroplach wody w oleju, aby naśladować przegrodkę wielkości komórki. Użyto fluorescencyjnego układu chemicznego do śledzenia, jak szybko spalane jest ATP, a jednocześnie obrazowano ruch i kurczenie się sieci aktynowej. Śledząc jednocześnie zużycie paliwa i ruch, oszacowano moc mechaniczną wytwarzaną przez miozynę oraz wydajność, z jaką energia była przekształcana w pracę.
Więcej aktyny, większa moc
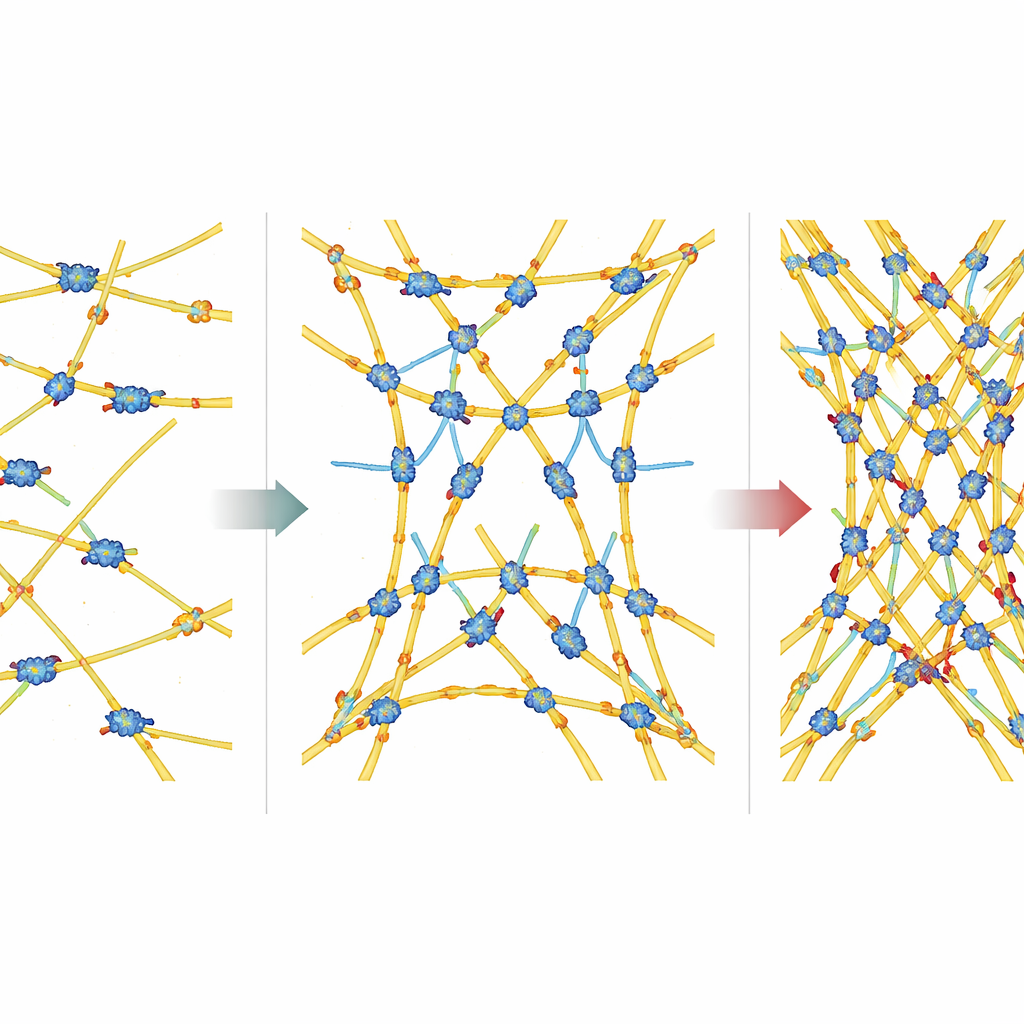
Pierwszym krokiem było sprawdzenie, jak zmiana ilości aktyny wpływa na motory. Wraz ze wzrostem stężenia aktyny miozyna spalała ATP szybciej, a sieć kurczyła się silniej. Zależność ta odpowiadała klasycznemu zachowaniu enzymatycznemu, wskazując, że motory zachowują się jak katalizatory, których prędkość zależy od ilości filamentów, do których mogą się przyłączyć. Na podstawie analizy ruchu na obrazach zespół obliczył „pozorny odkształcenie” (o ile zmniejszyła się objętość sieci) oraz „wnioskowaną moc mechaniczną” z tego skurczu. Obie wielkości rosły wraz ze stężeniem aktyny, podobnie jak pozorna wydajność — ułamek energii chemicznej, który wydawał się być zamieniany na pracę mechaniczną. Przy wysokich poziomach aktyny sieć stawała się sztywniejsza, co pozwalało na skuteczniejsze rozprzestrzenianie sił i zwiększało całkowitą moc wyjściową.
Wiązacze jako ukryte pokrętła regulacji
Komórki nie polegają wyłącznie na aktynie; używają białek krzyżujących, aby łączyć filamenty w różne architektury. Autorzy przetestowali cztery powszechne wiązacze — α-aktyninę, fascynę, fimbrynę i filaminę — które tworzą sieci o odmiennym rozstawie i wyrównaniu filamentów. Dodanie α-aktyniny, która tworzy stosunkowo luźne połączenia o mieszanej polaryzacji, zwiększyło zarówno zużycie ATP, jak i moc mechaniczną, ale wydajność pozostała w przybliżeniu taka sama. W przeciwieństwie do tego fascina, która spaja filamenty w ciasno upakowane, jednolicie wyrównane pęczki, spowalniała spalanie ATP przez miozynę, przy jednoczesnym zwiększeniu mocy mechanicznej i wydajności. Fimbryna i filamina wykazywały bardziej złożone zachowanie: przy niskich stężeniach podnosiły zużycie paliwa i moc, ale przy wyższych poziomach gwałtownie redukowały zużycie ATP, w niektórych przypadkach niemal zatrzymując motory, podczas gdy część sieci nadal była zdolna do kurczenia się.
Jak obciążenie zmienia sposób pracy motorów
Te wzorce wskazują na istotną ideę: motory miozynowe są wrażliwe na obciążenie mechaniczne. Gdy ciągną przeciwko sztywnej, silnie skrzyżowanej sieci, pozostają dłużej w stanie generującym siłę, skutecznie mocniej chwytając filament. Autorzy wykorzystali tempo zużycia ATP do oszacowania efektywnego „współczynnika pracy” (duty ratio), czyli ułamka czasu, w którym motor spędza w silnie związanym stanie, i połączyli to z mocą mechaniczną, aby ocenić, jak bardzo układ zależy od obciążenia. Sieci oparte na fascinie i filaminie wykazywały silną zależność od obciążenia: wraz ze wzrostem krzyżowania motory spowalniały cykl, ale dostarczały większą siłę w czasie. Sieci tworzone przez α-aktyninę i fimbrynę dawały bardziej umiarkowane zmiany. Te różnice odzwierciedlają miejsca występowania wiązaczy w komórkach — fascina i filamina w wysoce dynamicznych strukturach, które pchają i ciągną otoczenie, α-aktynina i fimbryna w bardziej stabilnych, wspierających rejonach.
Dlaczego to ma znaczenie dla żywych komórek
Mówiąc prosto, praca ta pokazuje, że komórki mogą stroić swoją „gospodarkę paliwową” i moc wyjściową nie tylko poprzez zmianę liczby używanych motorów, lecz także przez reorganizację rusztowania aktynowego, za które ciągną. Niektóre wiązacze tworzą architektury sprzyjające energetycznemu, wrażliwemu na obciążenie ciągnięciu, odpowiednie do szybkiego ruchu i przekształceń. Inne wspierają stabilne, mniej wrażliwe na obciążenie zachowanie, idealne do utrzymania struktury. Poprzez dobór i mieszanie tych wiązaczy komórki mogą dostosować, ile energii wydatkują na generowanie sił potrzebnych do podziału, migracji i rozwoju. Badanie to łączy zatem mikroskopowe detale architektury cytoszkieletu z szerszym pytaniem, jak systemy żywe zarządzają i wykorzystują swoją energię.
Cytowanie: Sakamoto, R., Sun, Z.G. & Murrell, M.P. Crosslinked F-actin networks regulate load-dependent energy conversion. Commun Biol 9, 572 (2026). https://doi.org/10.1038/s42003-026-09843-0
Słowa kluczowe: actomyosyna, cytoszkielet, mechanika komórkowa, zużycie ATP, wiązacze aktyny