Clear Sky Science · pl
Tworzenie klastrów rezystosomu pomocniczych NLR po aktywacji skręconej‑cewkowej NLR
Jak rośliny zamieniają wykrycie patogenu w komórkowe samobójstwo
Rośliny nie mogą uciec przed zarazkami, dlatego każda komórka ma własny system awaryjnej obrony. Jedną z najbardziej dramatycznych reakcji jest kontrolowana forma samobójstwa komórkowego, która poświęca zainfekowane komórki, by ocalić resztę rośliny. Artykuł ten ujawnia, jak konkretny receptor odpornościowy, zwany SUMM2, zlokalizowany przy powierzchni komórki, zbiera zespół białek pomocniczych w uderzające, pierścieniowe skupiska, które mogą przebijać zewnętrzną błonę komórkową, wymuszając tę decyzję o życiu lub śmierci.
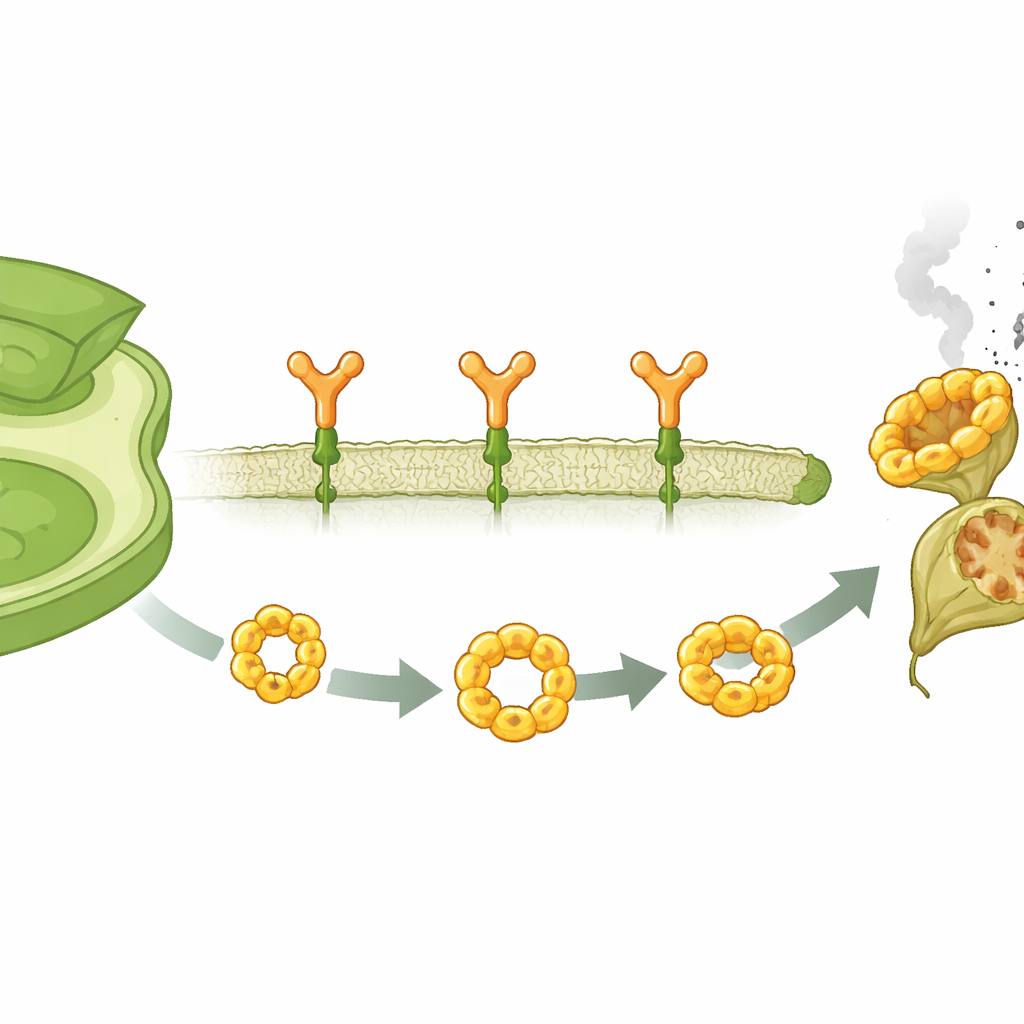
Strażnik przy zewnętrznej granicy komórki
Komórki roślinne otacza błona plazmatyczna, która pełni rolę linii frontu przeciwko najeźdźcom. Wewnątrz tej granicy znajduje się wiele receptorów odpornościowych monitorujących oznaki ataku. Badanie koncentruje się na jednym z takich receptorów, SUMM2, występującym w modelowej roślinie Arabidopsis. W przeciwieństwie do lepiej znanych receptorów, które wciskają się w błonę, tworząc proste pory, SUMM2 jest przytwierdzony do błony za pomocą małego tłuszczowego ogonka, dodanego przez modyfikację chemiczną zwaną N‑myrystylacją. Autorzy pokazują, że ten lipidowy znacznik jest powszechny w jednej klasie receptorów odpornościowych i jest niezbędny do umieszczenia SUMM2 w błonie oraz utrzymania jego stabilności tam. Gdy znacznik zostaje odjęty, SUMM2 przemieszcza się do wnętrza komórki, staje się mniej liczny i nie jest już w stanie skutecznie wywołać śmierci komórkowej.
Zawiadamianie wyspecjalizowanej drużyny ratunkowej
SUMM2 nie działa samodzielnie. Zespół badawczy odkrył, że gdy SUMM2 zostaje aktywowany — albo przez genetyczne sztuczki, albo przez bakteryjne białko efektorowe, które unieszkodliwia odrębny szlak sygnalizacyjny — rekrutuje trio białek pomocniczych znanych jako EDS1, PAD4 i ADR1. Te pomocnicze białka są już znane z roli w odporności roślin, lecz uważano, że działają głównie z inną klasą sensorów odpornościowych. Łącząc genetykę i testy interakcji białek, badacze wykazują, że rośliny pozbawione EDS1, PAD4 lub ADR1 w dużej mierze tracą zdolność do zachodzenia śmierci komórkowej indukowanej przez SUMM2 i wykazują łagodniejsze objawy związane z chorobą. Umieszcza to moduł EDS1–PAD4–ADR1 wyraźnie poniżej SUMM2 jako niezbędne ogniwo przekazujące początkowy alarm w pełną reakcję obronną.
Z ruchomych pomocników do zamrożonych pierścieni
Aby dowiedzieć się, co naprawdę dzieje się przy błonie, autorzy zastosowali obrazowanie w wysokiej rozdzielczości na żywych komórkach. W spoczynkowych komórkach cząsteczki ADR1 szybko przesuwają się po wewnętrznej powierzchni błony jako maleńkie, ruchome kropki. Po aktywacji SUMM2 to zachowanie zmienia się dramatycznie. Obszary z ADR1 zwalniają, stają się nieruchome i łączą się w małe grupy po dwie do sześciu jednostek ułożonych w porządne, pierścieniowe wzory osadzone w błonie. Te skupiska pojawiają się zanim wystąpią oczywiste oznaki śmierci komórkowej, co sugeruje, że są częścią mechanizmu wyzwalającego, a nie jedynie produktem ubocznym. Podobne zachowanie skupisk zaobserwowano dla pokrewnych receptorów pomocniczych w innych gatunkach roślin, co sugeruje, że takie zespoły mogą być powszechną cechą sygnalizacji odpornościowej roślin.
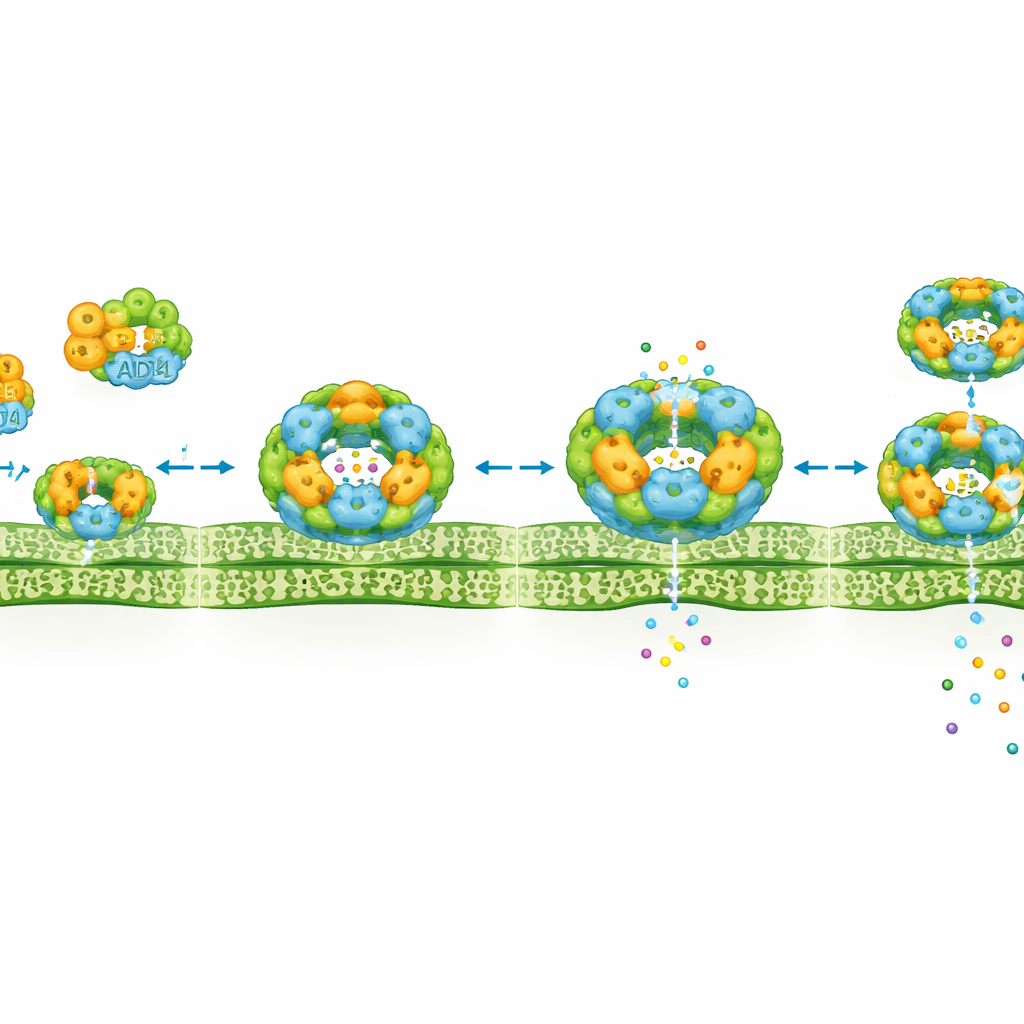
Budowanie maszyn śmierci wyższej rangi
Historia staje się jeszcze bardziej złożona, gdy EDS1 i PAD4 obserwuje się równolegle z ADR1. Samodzielnie EDS1 i PAD4 przemieszczają się przez jądro i cytoplazmę. Po aktywacji SUMM2 są przyciągane do błony i gromadzą się w tych samych pierścieniowatych miejscach, gdzie zgrupował się ADR1. Szczegółowe obrazy pokazują, że EDS1–PAD4 tworzą ciągły pierścień, podczas gdy liczne zespoły ADR1 punktują ten pierścień niczym koraliki. Doświadczenia biochemiczne potwierdzają ten obraz, ujawniając, że aktywacja SUMM2 sprzyja tworzeniu dużych kompleksów zawierających razem EDS1, PAD4 i ADR1. Chemiczne inhibitory blokujące produkcję małych cząsteczek sygnałowych wytwarzanych przez jeszcze inną grupę białek odpornościowych uniemożliwiają zarówno formowanie skupisk, jak i śmierć komórkową, co sugeruje, że te skupiska integrują sygnały z kilku warstw sieci obronnej roślin.
Dlaczego te pierścienie mogą być śmiertelne
Co tak naprawdę robią te uderzające struktury? Wcześniejsze prace wykazały, że pokrewne kompleksy odpornościowe mogą tworzyć małe kanały, które wpuszczają skoki jonów wapnia do komórki — kluczowy krok w sygnalizacji obronnej. Nowe badanie sugeruje, że SUMM2 używa swojej lipidowej kotwicy nie do tworzenia własnych porów, ale do aranżowania skupisk kompleksów pomocniczych w większe pierścienie, które mogą silniej zaburzać błonę. Autorzy wysuwają przypuszczenie, że, podobnie jak niektóre białka w układach odpornościowych zwierząt, które wycinają fragmenty błony, by zniszczyć umierające komórki, te roślinne pierścieniowe zespoły mogłyby lokalnie osłabić lub usunąć kawałki błony, pozwalając na wyciek zawartości i przypieczętowując los komórki. Mówiąc prościej: SUMM2 działa jak przytwierdzony strażnik, który, gdy zostaje przekonany o obecności patogenu, zbiera ekipę demolującą na powierzchni komórki i rozkazuje im tworzyć zorganizowane otwory, które zamieniają pojedynczą zainfekowaną komórkę w ochronną ofiarę dla rośliny.
Cytowanie: Ge, D., Ortiz-Morea, F.A., Xie, Y. et al. Assembly of helper NLR resistosome clusters upon activation of a coiled-coil NLR. Nature 652, 251–258 (2026). https://doi.org/10.1038/s41586-026-10215-1
Słowa kluczowe: odporność roślin, śmierć komórkowa, receptory odpornościowe, skupiska białek błonowych, Arabidopsis