Clear Sky Science · pl
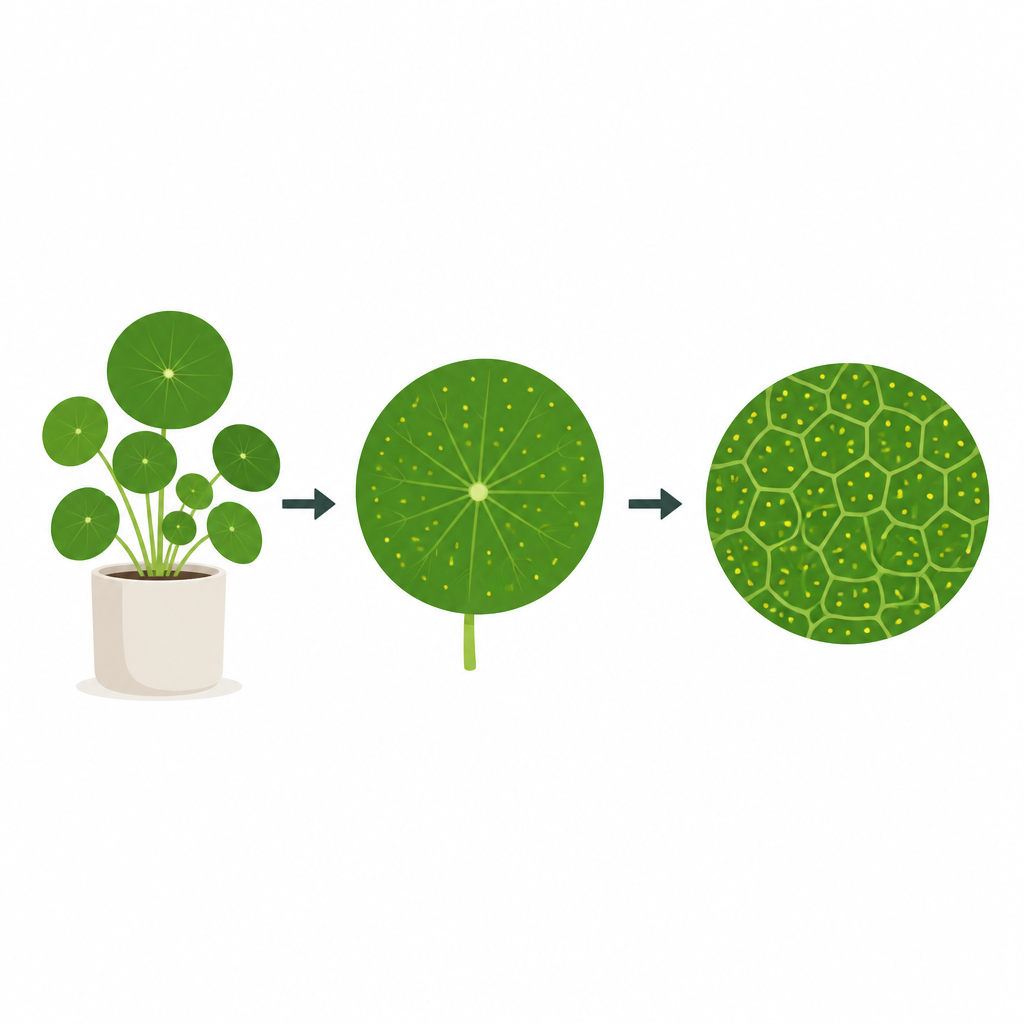
Siatkowata wiązkowatość liści Pilea peperomioides to diagram Voronoi
Jak roślina doniczkowa ukrywa zagadkę matematyczną
Wiele osób hoduje chińską monetę dla jej okrągłych, monetopodobnych liści, ale niewielu zgadłoby, że te liście cicho stosują regułę z lekcji geometrii. Badanie pokazuje, że największe żyły w tych liściach układają się podobnie do klasycznej łamigłówki zwanej diagramem Voronoi, czyli sposobu podziału przestrzeni na obszary wokół określonych punktów. Ujawniając zarówno wzór, jak i krok po kroku biologiczny mechanizm, który może go wytworzyć, praca łączy codzienne formy roślin z prostymi zasadami matematycznymi.
Dostrzeganie wzorów w żyłkowaniu liści
Autorzy koncentrują się na Pilea peperomioides, której niemal okrągłe liście są przymocowane do łodygi via ogonków z dołu. Każdy liść zawiera sieć grubych „głównych” żyłek tworzących zamknięte pętle oraz drobniejszą siatkę cienkich naczyń. Rozrzucone po powierzchni liścia są hydatody — maleńkie pory wydalające wodę i pomagające w regulacji równowagi wewnętrznej liścia. Gdy badacze zabarwili i zaobrazowali spłaszczone liście, a następnie komputerowo wyśledzili każdą główną żyłkę i każdy hydatod, zauważyli uderzający fakt: większość najmniejszych pętli głównych żyłek obejmowała dokładnie jeden hydatod. Sugerowało to, że żyłki mogą działać jak granice narysowane w połowie drogi między sąsiednimi porami.
Aby sprawdzić ten pomysł, sięgnęli do diagramów Voronoi, które dzielą przestrzeń na komórki wokół zbioru punktów tak, że każda lokalizacja należy do najbliższego punktu. Badacze porównali rzeczywiste pętle żyłek z idealnymi komórkami Voronoi zbudowanymi z pozycji hydatodów. Użyli trzech niezależnych testów geometrycznych: jeden sprawdzał, czy linie między sąsiednimi hydatodami przecinają wspólną krawędź żyłki pod kątem prostym i w równych odległościach; inny mierzył, ile powierzchni każda rzeczywista pętla dzieliła z odpowiadającą jej komórką Voronoi; trzeci działał wstecz, pytając, gdzie powinny leżeć najlepiej dopasowane centra Voronoi dla zaobserwowanej sieci i jak blisko tych centrów były rzeczywiste hydatody. We wszystkich testach hydatody konsekwentnie zachowywały się bardziej jak centra Voronoi niż kilka alternatywnych punktów odniesienia wewnątrz każdej pętli.
Wzory, które utrzymują się pod stresem
Wzrost biologiczny nigdy nie jest idealnie regularny, a liście mogą być przekształcane przez środowisko. Aby sprawdzić, jak odporne jest ułożenie podobne do Voronoi, zespół hodował rośliny w cieniu, przy intensywnym świetle i w wysokiej temperaturze, po czym przeanalizował ponad sto nowych liści. Te warunki zmieniały rozmiar liścia, kolor i rozmiar hydatodów, ale nie średnią liczbę hydatodów na liść ani ich ogólny rozkład przestrzenny. Co ważne, te same trzy testy geometryczne wykazały, że relacja między hydatodami a głównymi żyłkami pozostawała bliska idealnemu diagramowi Voronoi we wszystkich warunkach. Symulacje sugerowały, że obserwowane odchylenia można wyjaśnić przez dodanie tylko umiarkowanego losowego szumu do doskonałego diagramu. Ta stabilność wskazuje na lokalny, samoregulujący się mechanizm, a nie sztywny, z góry ustalony plan.
Fala chemiczna, która rysuje mapę
Następne pytanie brzmiało, jak żywe komórki mogłyby wygenerować taki wzór. Biolodzy roślin od dawna faworyzują ideę „kanałowania” (canalization), w której hormon wzrostu — auksyna — płynie od źródeł do odpływów i poprzez sprzężenie zwrotne z białkami transportującymi (znanymi jako PIN) wyrzeźbia kanały o wysokim przepływie, które stają się żyłami. Kanałowanie naturalnie tworzy rozgałęzione, drzewiaste ścieżki łączące źródła i odpływy, ale ma trudności z wyjaśnieniem zamkniętych pętli leżących między źródłami a nie łączących je. Autorzy proponują inną, opartą na auksynie mechanikę: hydatody zachowują się jak źródła auksyny, ale zamiast tworzyć bezpośrednie kanały, wysyłają rozchodzące się fale wysokiego stężenia auksyny. Tam, gdzie fale od sąsiednich hydatodów się zderzają, pojawiają się grzbiety podobne do grzebieni dokładnie między nimi, wyznaczając ostateczne ścieżki głównych żyłek.
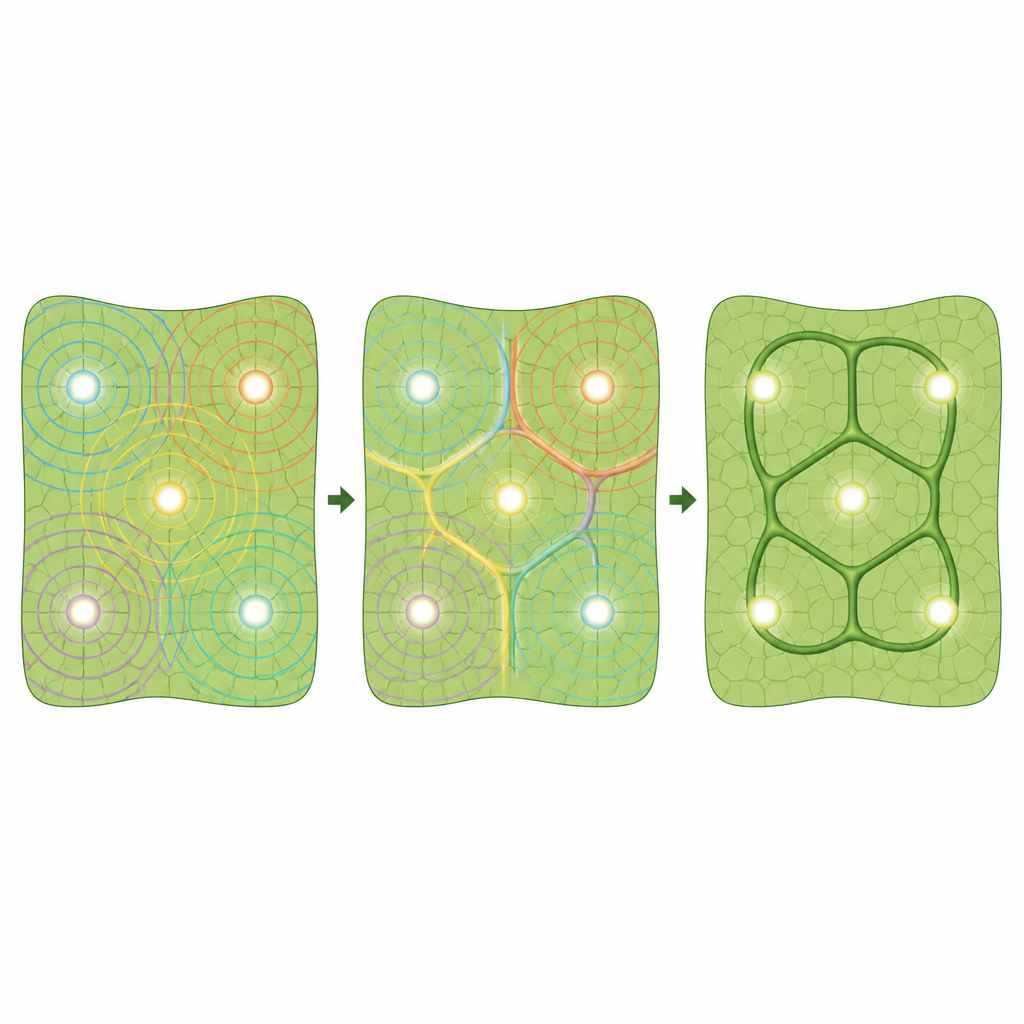
Od modelu do żywego liścia
Wykorzystując symulacje komputerowe siatki komórek, zespół pokazał, że gdy transport auksyny jest tylko słabo spolaryzowany w jednym kierunku, z każdego źródła wyłaniają się fale, przemieszczają się przez tkankę i budują grzbiety na liniach zderzeń. W dwuwymiarowej, o kształcie liścia siatce zainicjowanej rzeczywistymi pozycjami hydatodów te grzbiety tworzą pętle, które ściśle odpowiadają zarówno idealnemu diagramowi Voronoi, jak i rzeczywistym głównym żyłkom, szczególnie przy krawędzi liścia. Model dopracowano, dodając reguły dla momentu różnicowania komórek żyłkowych i tego, jak poziomy białek PIN odpowiadają na auksynę, co doprowadziło do lepszego dopasowania symulowanych wzorców PIN do obrazów mikroskopowych. Ponieważ narzędzia reporterowe genetyczne nie są jeszcze dostępne w Pilea, badacze użyli przeciwciał rozpoznających białka PIN, aby zmapować ich występowanie podczas rozwoju liścia. Znaleźli silny sygnał PIN wokół hydatodów i w żyłkach pierwotnych, niewielki sygnał w samych żyłkach drugorzędnych oraz spolaryzowany PIN w komórkach sąsiednich wskazujący w stronę tych żyłek, co zgadza się z ideą fal auksyny kształtujących sieć z źródeł skupionych przy hydatodach.
Dlaczego to ma znaczenie poza jedną rośliną doniczkową
Mówiąc prosto, badanie dochodzi do wniosku, że chińska moneta rysuje swoje pętliste żyłki liściowe za pomocą reguły geometrycznej, w której każdy hydatod rości sobie „terytorium”, a granice między terytoriami stają się głównymi żyłkami. Reguła ta może być wytworzona przez chemiczne fale auksyny rozchodzące się z wielu punktów i oznaczające swoje granice w momencie spotkania. Ponieważ podobne pory i układy żyłek występują u innych gatunków, ten sam mechanizm „fala-i-granica” może pomóc wyjaśnić szeroką gamę sieciowych wzorów żyłkowych roślin. Szerzej rzecz biorąc, wyniki pokazują, jak tkanki żywe mogą wykorzystywać proste reguły odległości i równoważenia sił do budowania złożonych struktur, które wyglądają skomplikowanie, ale mają podłoże w prostych zasadach matematycznych.
Cytowanie: Zheng, C.X., Palit, S., Venezia, M. et al. Reticulate leaf venation in Pilea peperomioides is a Voronoi diagram. Nat Commun 17, 4111 (2026). https://doi.org/10.1038/s41467-026-71768-3
Słowa kluczowe: wiązkowatość liścia, wzór Voronoi, fale auksyny, geometria roślin, Pilea peperomioides