Clear Sky Science · pl
Strukturalne i dynamiczne wglądy w rozpoznawanie agonistów i funkcję receptora tromboksanu A2
Dlaczego ten strażnik krzepnięcia krwi ma znaczenie
Organizm utrzymuje płynny przepływ krwi, zachowując jednocześnie zdolność do zatamowania krwawienia w kilka sekund. Kluczowym elementem tej równowagi jest receptor tromboksanu A2, niewielki przełącznik białkowy na płytkach krwi i komórkach mięśni gładkich naczyń, który reguluje krzepnięcie, zwężenie naczyń i stan zapalny. Gdy ten przełącznik zawodzi lub jest nadmiernie aktywny, może przyczyniać się do zawałów serca, udarów, chorób płuc, włóknienia, a nawet niektórych nowotworów. Niniejsze badanie ujawnia w bezprecedensowych, atomowych szczegółach, jak receptor rozpoznaje swoje aktywujące cząsteczki i jak ulega aktywacji wewnątrz komórki — wiedza ta może ukierunkować bezpieczniejsze i precyzyjniejsze leki do regulacji krzepnięcia i napięcia naczyniowego.
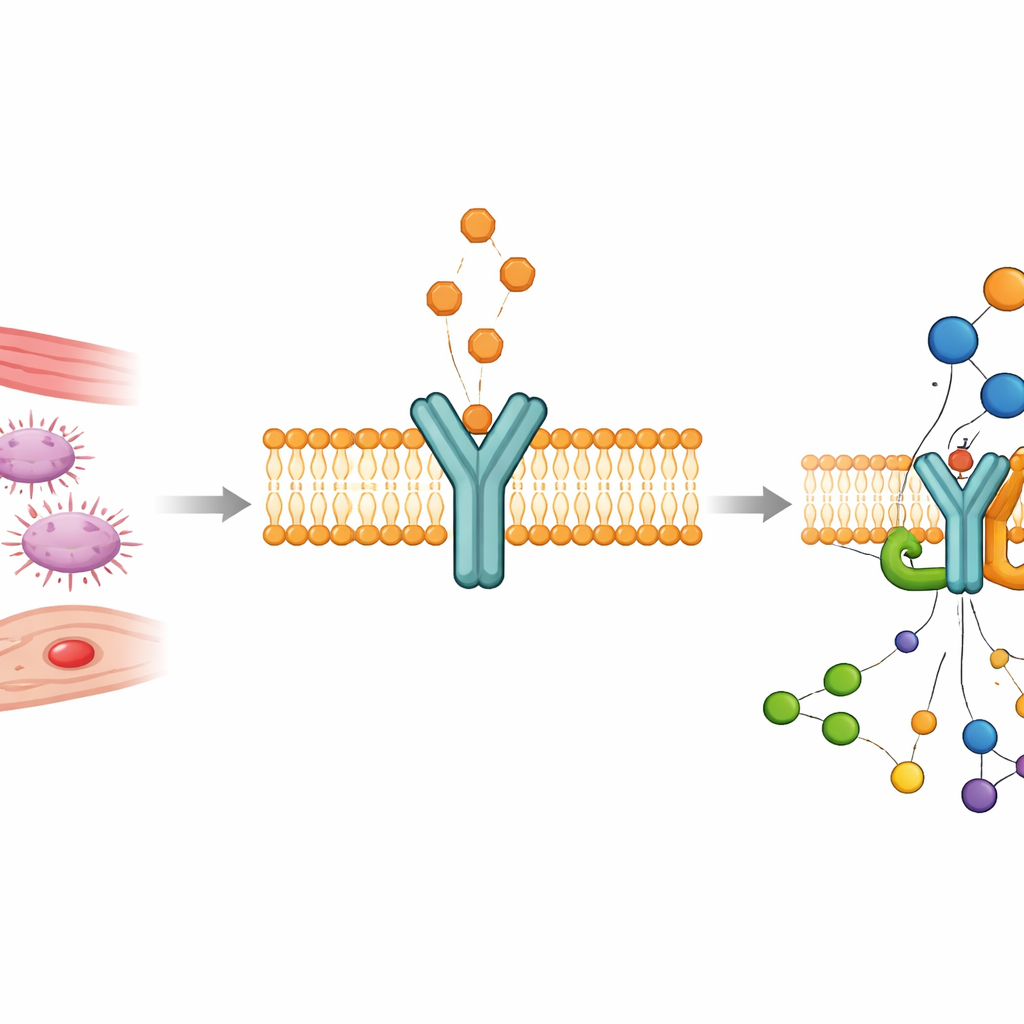
Krótkotrwały sygnał o długotrwałych skutkach
Tromboksan A2 jest pochodzącym z tłuszczów przekaźnikiem produkowanym w płytkach krwi i ścianach naczyń. Jest chemicznie nietrwały, rozkłada się w około pół minuty, lecz w tym krótkim oknie instruuje płytki, by się zbijały, i powoduje kurczenie się otaczających mięśni. Robi to, wiążąc się z receptorem tromboksanu A2 — członkiem dużej rodziny receptorów sprzężonych z białkiem G, osadzonych w błonie komórkowej, przekazujących sygnały do wnętrza komórki. Ponieważ naturalny tromboksan tak szybko znika, trudno było uchwycić, w jaki sposób dokuje do receptora i przełącza go w stan aktywny. Badacze ominęli to ograniczenie, używając dwóch stabilnych, wysokoczynnych substytutów, U46619 i I‑BOP, które naśladują tromboksan, lecz utrzymują się wystarczająco długo, by móc je zbadać strukturalnie.
Obserwacja receptora w akcji
Przy użyciu krioelektronowej mikroskopii zespół uwiecznił ludzki receptor związany z każdym z tych syntetycznych agonistów, podczas gdy był sprzężony ze swoim głównym wewnątrzkomórkowym partnerem, białkiem Gq. Powstałe trójwymiarowe struktury, wyostrzone do niemal atomowej rozdzielczości, pokazują receptor przecinający błonę komórkową siedmioma ciasno upakowanymi helisami. Cząsteczki agonistów siedzą głęboko wewnątrz tego pęku w pozycji przypominającej literę L, chronione przed wodą, która w przeciwnym razie mogłaby je zniszczyć. Po wewnętrznej stronie komórki jedna z helis odchyla się na zewnątrz, otwierając zagłębienie, w które wsuwa się ogon białka G, przygotowując komórkę na napływ następnych sygnałów wapniowych i aktywację enzymów. Biochemiczne testy sygnałowe potwierdziły, że te kompleksy wiernie odzwierciedlają aktywny, funkcjonujący stan receptora.
Ukryte przejście przez błonę
Struktury wraz z rozległymi symulacjami komputerowymi wskazują, że cząsteczki podobne do tromboksanu nie podchodzą głównie do receptora od wodnego zewnętrza komórki. Zamiast tego wsuwają się z wnętrza tłuszczowej warstwy błony. Ruchoma szczelina między dwiema z zewnętrznych helis, wspomagana przez elastyczne aromatyczne łańcuchy boczne i pobliskie cząsteczki cholesterolu, działa jak bramka molekularna. W stanie nieaktywnym szczelina jest otwarta, umożliwiając wejście; gdy agonista jest już zakopany w kieszeni, bramka zamyka się, uszczelniając ligand na miejscu. Mutacje kluczowych reszt tworzących tę bramkę lub zmiany ich interakcji z cholesterolem poważnie zakłócają sygnalizację receptora — co pomaga wyjaśnić rzadkie ludzkie zaburzenia krwawienia spowodowane naturalnie występującymi mutacjami w tych miejscach.
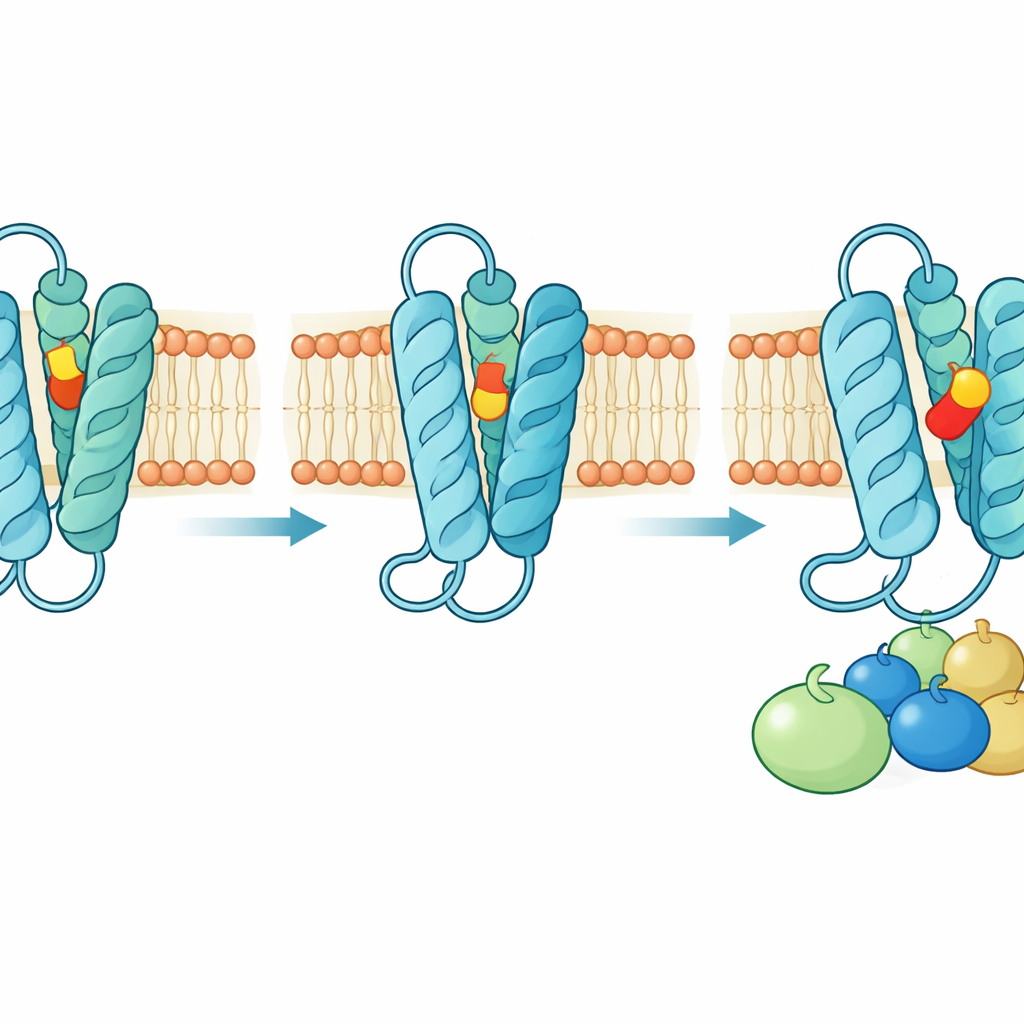
Niekonwencjonalny sposób przełączania
Większość powiązanych receptorów aktywuje się, gdy ligand popycha konserwatywny aminokwas działający jak „przełącznik”, co z kolei powoduje duży ruch jednej z helis. W receptorze tromboksanowym ten klasyczny toggle istnieje, lecz pełni rolę wspierającą. Zamiast niego badanie wykazuje, że para reszt glutaminy i asparaginy położona po wewnętrznej stronie kieszeni ligandowej działa jako prawdziwy przełącznik. Agoniści przekazują wiązanie wodorowe w sposób, który zmusza te reszty do przegrupowania swoich połączeń, przyciągając jedną helisę do środka, pozwalając innej na rotację i ostatecznie uwalniając helisę wychodzącą na zewnątrz, która tworzy miejsce dla białka G. Antagonisty natomiast siedzą wyżej w kieszeni i wiążą się inaczej z tą samą glutaminą, stabilizując nieaktywną sieć kontaktów, która zamyka miejsce wiązania białka G. Dziesiątki starannie dobranych mutacji, testowanych w żywych komórkach, potwierdzają tę zrewidowaną koncepcję mechanizmu włączania i wyłączania receptora.
Od atomowych planów do lepszych leków
W sumie praca dostarcza pełnego mechanistycznego obrazu, w jaki sposób nietrwałe sygnały tromboksanu są przechwytywane z błony, zamykane w osłoniętej wnęce i przekształcane w silną odpowiedź wewnątrzkomórkową za pośrednictwem nietypowego mechanizmu aktywacji. Wyjaśniając, dlaczego niektóre ligandy silnie aktywują receptor, a inne go blokują, oraz mapując, jak mutacje związane z chorobami wypaczają te procesy, badanie oferuje szczegółowy plan dla leków następnej generacji. Takie związki mogłyby precyzyjniej tłumić nadmierne krzepnięcie, skurcz naczyń i włóknienie — lub selektywnie blokować szkodliwe sygnały tromboksanu w nowotworach — przy jednoczesnym zachowaniu istotnej roli receptora w codziennym zdrowiu naczyń.
Cytowanie: Krawinski, P., Matzov, D., Ryder, A. et al. Structural and dynamic insights into agonist recognition and function of the thromboxane A2 receptor. Nat Commun 17, 3071 (2026). https://doi.org/10.1038/s41467-026-69844-9
Słowa kluczowe: receptor tromboksanu, krzepnięcie krwi, receptor sprzężony z białkiem G, struktura cryo-EM, choroba układu krążenia