Clear Sky Science · nl
Structurele organisatie van het HBV pgRNA-genoom gestuurd door fasescheiding in capsidebeperking
Hoe een piepklein virus een groot genoom inpakt
Het hepatitis B-virus (HBV) is een belangrijke oorzaak van leverziekten wereldwijd, en toch past zijn genetisch materiaal in een eiwitschil van slechts ongeveer een honderdmiljardste meter breed. Dit artikel onderzoekt een eenvoudig maar al lang bestaand raadsel: hoe kan het virus zijn RNA-genoom in zo'n nauwe ruimte proppen terwijl het nog beweeglijk genoeg blijft om zichzelf te kopiëren? Met behulp van computersimulaties en laboratoriumexperimenten onthullen de auteurs een fysisch proces, vergelijkbaar met hoe oliedruppels zich in water vormen, dat HBV in staat stelt zijn genoom netjes in de schaal te organiseren en mogelijk nieuwe manieren biedt om het virus te verstoren.
Een drukke wereld in de virale schaal
In de eiwitschil van HBV, de capsid, bevindt zich het pregenomische RNA (pgRNA), een lange enkelstrengseltje dat dient als sjabloon voor het maken van viraal DNA. Het binnenoppervlak van de schaal is bezaaid met flexibele, positief geladen eiwitstaartjes die aangetrokken worden door het negatief geladen RNA. Gedetailleerde atomistische simulaties laten zien dat het pgRNA, in plaats van een vaste klomp in het midden te vormen, zich snel naar de binnenwand verplaatst en daar een holle, schaalachtige laag vormt die dicht tegen de capsid aanligt. Binnen deze laag bestaan dichte patches van RNA en eiwitstaartjes naast meer open, poreuze regio's. Gemiddeld komt deze ordening overeen met de sterk symmetrische patronen die zichtbaar zijn in cryo-elektronenmicroscopiebeelden, maar elk afzonderlijk virusdeeltje kan op een gegeven moment behoorlijk verschillend ogen. 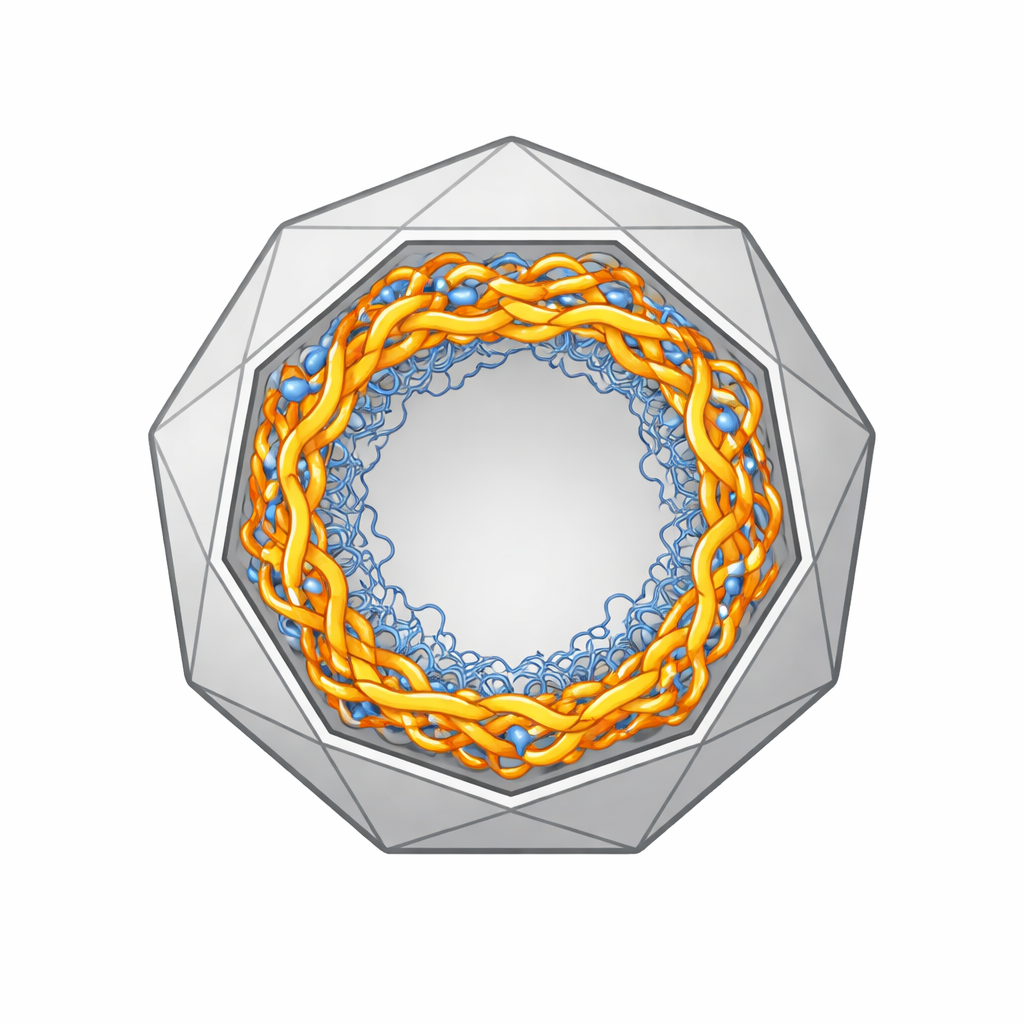
Druppels zonder container
Om te begrijpen wat dit patroon aandrijft, gingen de onderzoekers over op grovere, snellere simulaties en aanvullende reageerbuisexperimenten. Ze vonden dat het RNA en de eiwitstaartjes een soort microscopische demenging ondergaan die bekend staat als vloeistof–vloeistof fasescheiding: ze vormen dichte, druppelachtige condensaten die naast meer verdunde omgevingen bestaan. Bij lage zoutconcentraties of lage temperaturen is de elektrostatische aantrekkingskracht tussen de positief geladen staartjes en het negatief geladen RNA sterk, en worden de condensaten uitgesprokener en meer gevlekt. Het verhogen van zout of temperatuur verzwakt deze aantrekkingen en maakt de RNA-laag uniformer. Vergelijkbaar gedrag treedt zelfs op wanneer de staartjes aan een vlak oppervlak verankerd zijn in plaats van aan een gekromde capsid, en wanneer ze worden gemengd met korte RNA-fragmenten in bulkoplossing, wat ondersteunt dat deze neiging tot fasescheiding een intrinsieke eigenschap van het mengsel is.
Verborgen orde in een flexibel genoom
De dichte patches die door dit proces ontstaan doen meer dan alleen moleculen bijeenbrengen. Binnen die patches vouwt het RNA vaker terug op zichzelf en vormt korte dubbelstrengssegmenten en haarspeldstructuren, terwijl nabijgelegen regio's flexibel als enkelstrengen blijven. Simulaties tonen aan dat onder condities die fasescheiding bevorderen, het aantal base-gepaarde segmenten scherp toeneemt, en dat veel van deze dubbelstrengse stukken parallel georiënteerd raken en geordende rijen vormen. Deze geordende "eilanden" zijn met elkaar verbonden door zachtere, mobielere enkelstrengs schakels, waardoor het genoom een boomachtige architectuur krijgt die compact maar niet star is. Wanneer de auteurs de geladen staartjes verstoren, ze losknippen van de schaal of hun lading neutraliseren, nemen zowel de fasescheiding als het basepairen sterk af. Dit wijst erop dat het binnenoppervlak van de capsid, via zijn verankerde staartjes, actief de hogere orde van het genoom vormgeeft.
De virale kopieermachine in beweging houden
HBV moet zijn RNA-genoom in DNA omzetten binnen de afgesloten capsid, een proces uitgevoerd door een viraal enzym genaamd polymerase. Tijdens deze omzetting moet de polymerase tussen verre plaatsen langs het RNA springen—bewegingen die afhangen van langafstand-baseparing en van het vermogen van het enzym om zich door het inwendige te verplaatsen. Wanneer polymerase aan de simulaties wordt toegevoegd, creëert de fasescheiden-deurende, holle-schaalordening van het RNA open kanalen in het midden van de capsid waar het enzym de voorkeur geeft te verblijven en snel te diffunderen. Tegelijkertijd ondersteunt de geordende RNA-structuur meer langafstand base-pairing contacten, waarvan wordt gedacht dat ze de sjabloonsprongen van de polymerase sturen. Als fasescheiding wordt onderdrukt door de eiwitstaartjes te neutraliseren, vult het RNA het binnenste meer uniform, wikkelt het zich strak rond de polymerase en vertraagt diens beweging. 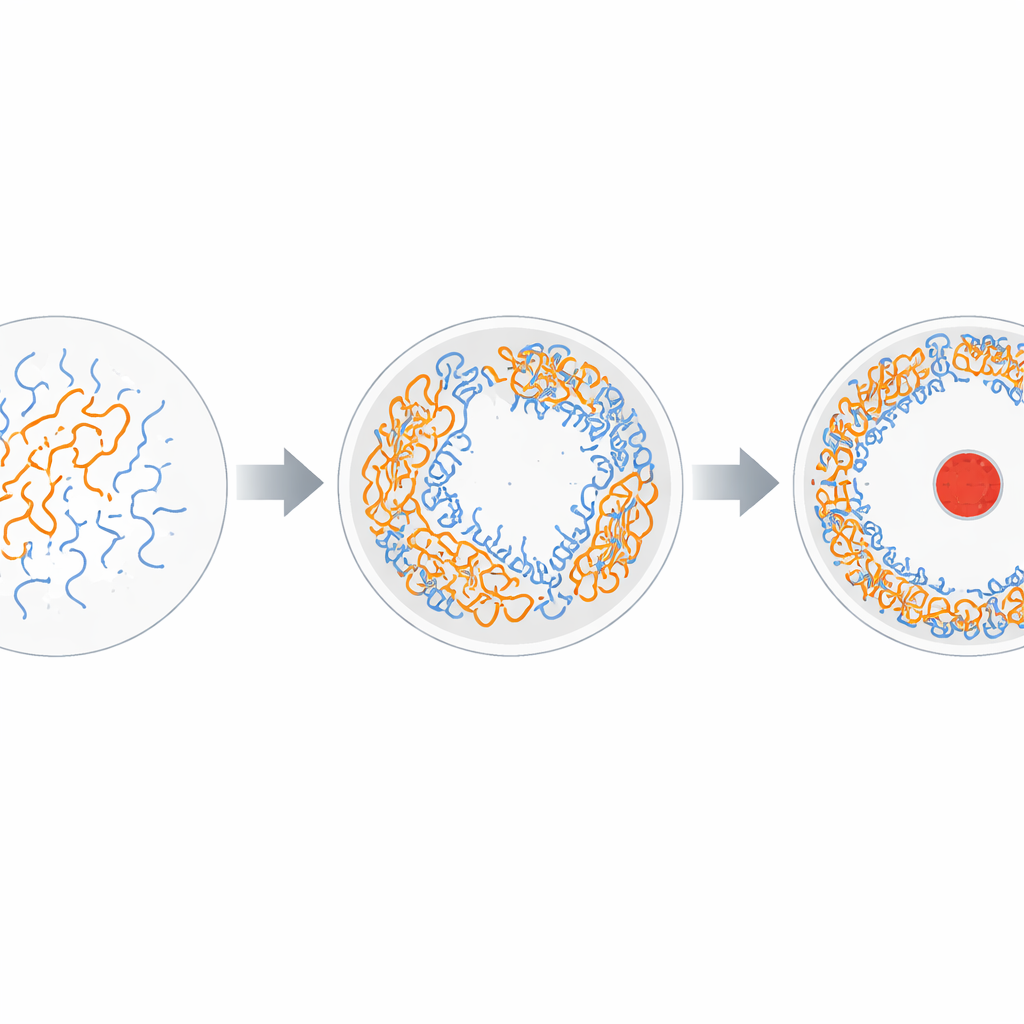
Waarom dit ertoe doet bij de behandeling van hepatitis B
Samen suggereren deze resultaten dat HBV een fundamenteel natuurkundig principe—fasescheiding—gebruikt om een ontwerpprobleem op te lossen: hoe een lang genoom in een kleine schaal te passen terwijl het ordelijk genoeg blijft voor nauwkeurige kopieën en tegelijkertijd los genoeg voor enzymen om te bewegen. Het virus bereikt dit door een holle laag van RNA–eiwitcondensaat langs de schaalwand te vormen, doorspekt met microdomeinen van geordende haarspelden en flexibele schakels, en door een opener binnenruimte voor de polymerase over te laten. Omdat deze organisatie sterk afhankelijk is van ladingsbalans en zoutcondities, zou het mogelijk zijn geneesmiddelen of peptiden te ontwerpen die de vorming of stabiliteit van het condensaat verstoren. Het richten op deze fysische laag van genoomorganisatie zou een nieuw pad naar antivirale therapieën kunnen bieden die complementair zijn aan benaderingen die direct gericht zijn op virale enzymen of instapstappen.
Bronvermelding: Bian, Y., Pan, H., Mao, J. et al. Structural organization of HBV pgRNA genome driven by phase separation in capsid confinement. Nat Commun 17, 2940 (2026). https://doi.org/10.1038/s41467-026-69689-2
Trefwoorden: hepatitis B-virus, organisatie van viraal genoom, vloeistof-vloeistof fasescheiding, RNA-condensaten, capsidestructuur