Clear Sky Science · ja
B2由来の抑制リボザイムの構造アーキテクチャを組織する切断領域
DNAに潜む隠れたスイッチ
私たちのDNAの多くは、かつてジャンクと片付けられた反復配列で満たされています。本研究はマウスに存在するそのような配列の一つ、B2に着目し、それが小さな分子スイッチのように振る舞い、自らを切断して細胞がストレス下にあるときに遺伝子の活動を停止するのに寄与することを示します。このスイッチがどのように組み立てられ、三次元的にどのように動くかを理解することは、発生、感染、がん、神経疾患との関連を含め、細胞が厳しい条件下で迅速に遺伝子をオフにする仕組みを説明する助けになります。 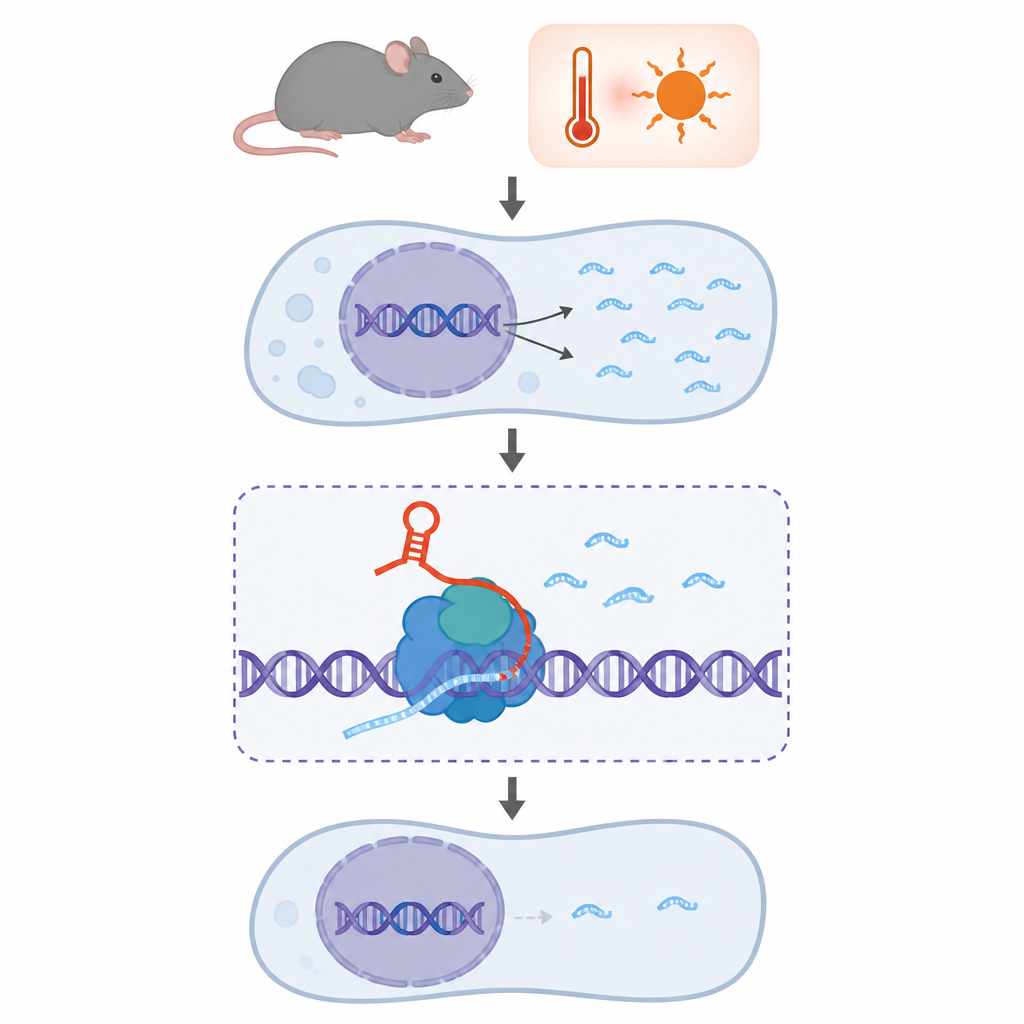
忙しい遺伝子を鎮める反復RNA
B2は哺乳類ゲノムに散在する短い反復要素のファミリーに由来します。マウスでは、B2 RNAは細胞の酵素によってDNAから転写され、初期胚や熱ショックやウイルス感染のようなストレス時に特に活性化されます。これまでの研究で、B2 RNAは主要な遺伝子読み取り機であるRNAポリメラーゼIIに結合して多くの遺伝子の転写を遅らせたり遮断したりできることが示されていました。さらに最近、B2は自己切断リボザイムであることが判明し、その活性はクロマチン修飾で知られるタンパク質EZH2によって調節されることが分かりました。これによりB2は遺伝子制御、ストレス応答、エピジェネティクスの交差点に位置付けられます。
小さな分子ナイフの形作り
B2の形が機能を支える仕組みを解明するために、著者らは複数の手法を組み合わせました。化学プロービング(SHAPEと呼ばれる)はRNAのどの部分が硬くどの部分が柔らかいかを明らかにし、茎ループといった二次構造のパターンを描き出しました。小角X線散乱(SAXS)は溶液中のRNAの低解像度三次元輪郭を提供し、単一の剛直な形だけでなく形のアンサンブルを捉えました。コンピュータモデリングはこれらのデータを全原子モデルに縫い合わせ、特定の領域がどのように折りたたまれ、動き、相互作用するかを可視化しました。研究は、自己切断部位を含み、EZH2結合やRNAポリメラーゼIIの阻害に重要な中心的な「切断領域」に焦点を当てました。
重要領域を変えたら何が起きるか
研究チームは標準的なB2 RNAといくつかの自然変異および設計変異を比較しました。B2Jと呼ばれる自然型はわずか二つの点変異を持ちます。二次構造の大枠はほぼ保たれるものの、三次元的にはより柔軟になり、より多くのコンフォメーションをサンプリングして自己切断が弱まります。主要な切断部位だけを欠く変異体B2Δ(96–105)は、意外にも近傍の茎が再配列してより長くて堅いアームを形成します。全体の大きさはほぼ同じですが、触媒活性と転写抑制能の多くを失い、この領域の硬直化が活性形へのアクセスを制限することを示唆しています。 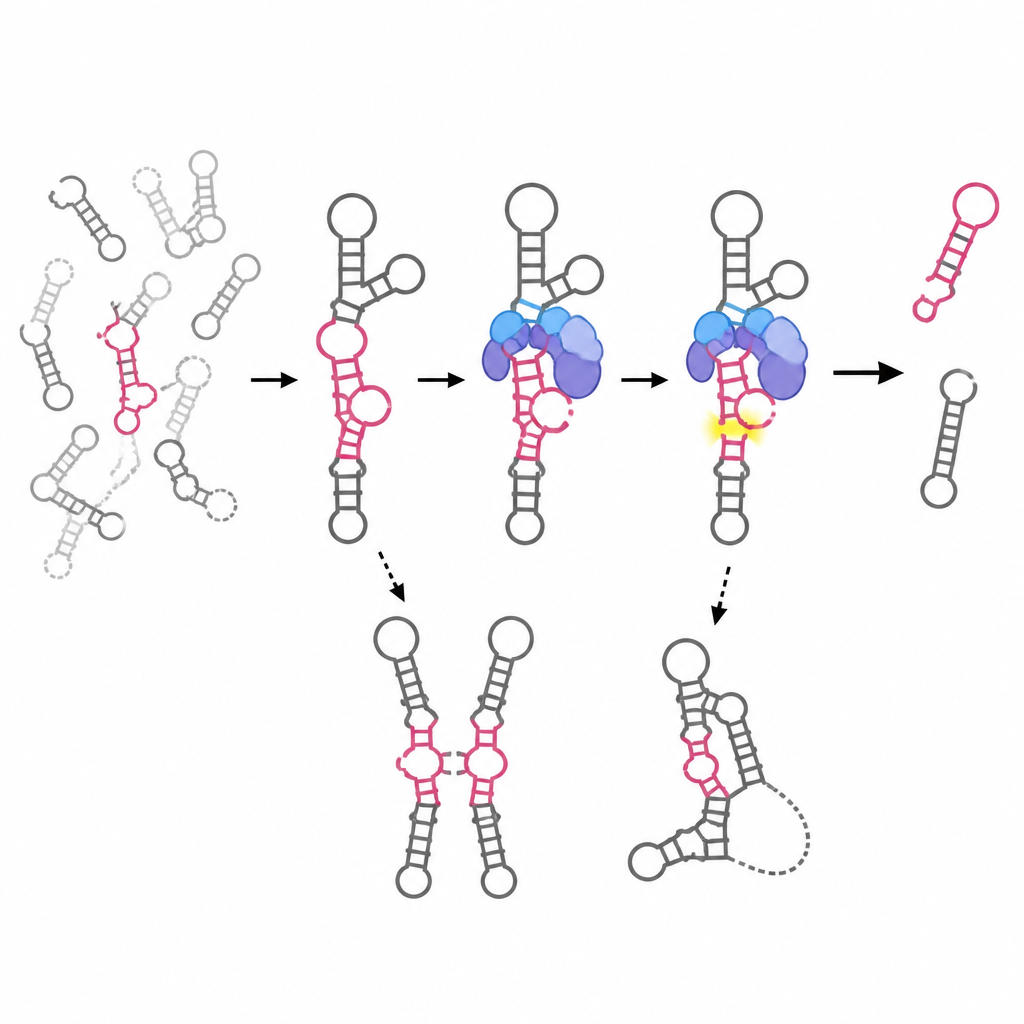
切除でどう変わるか
さらに大きな欠失であるB2Δ(81–124)は切断ドメイン全体を除去します。化学プロービングは、5′領域は保たれるものの残りの多くが構造を失うことを示します。SAXSはこの変異体がより大きく細長く見えることを示し、モデリングと結合試験は一本鎖のままではなくRNA二量体を形成できることを示唆します。このアーキテクチャの変化はEZH2結合の消失および転写抑制能の完全な喪失と一致し、試験管内の核抽出液を用いたアッセイでも、生細胞内でもこの変異体は通常のB2が示すような全体的なRNA合成の低下や核形態の変化を引き起こしません。
なぜこれらの可動部が重要か
総じて、本研究はB2 RNAの切断領域が単に切断部位であるだけでなく、その全体構造と機能を組織する要であることを示しています。微妙な点変化はRNAをより柔らかく効率を下げ、一方で領域を硬化させたり消失させたりする欠失はEZH2との協働やRNAポリメラーゼIIの抑制機能を損ないます。特定の三次元形状と形のアンサンブルを個別の生物学的結果に結び付けることで、いわゆるジャンクDNAの小片が精密に調整されたストレス応答性のブレーキとして機能し得る理由を説明しています。
引用: Singhal, A., Mrozowich, T., Rivera, C. et al. Cleavage region organizes the structural architecture of the SINE-derived B2 repressive ribozyme. Commun Biol 9, 649 (2026). https://doi.org/10.1038/s42003-026-09819-0
キーワード: B2 SINE RNA, 自己切断リボザイム, RNA構造, ストレス応答, 転写抑制