Clear Sky Science · ja
CM1活性化による微小管核生成中のヒトγTuRC閉鎖の構造基盤
細胞内の足場はどう作られるか
すべての動物細胞の内部には、染色体移動、貨物輸送、細胞形状の維持を助ける微細な管状の隠れた足場があります。本研究は、これらの管の形成を開始する主要な細胞機械が、どのように待機状態から活性状態へと反転するかを明らかにします。この微小なスイッチを理解することは、細胞が内部構造を確実かつ適時に維持する仕組みを知る手がかりになります。
微小管の出発プラットフォーム
微小管は、繰り返し配列するタンパク質サブユニットからできた中空の管で、通常13本の並列するプロトフィラメントで構成されます。この過程を始めることは困難であるため、細胞はγ-チュブリンリング複合体(γTuRC)と呼ばれる環状のプラットフォームに頼ります。この大きなタンパク質アセンブリは、最初のサブユニット列を整列させるテンプレートとして働きます。酵母のような単純な生物では、γTuRCは必要な場所、すなわち微小管ネットワークを組織する細胞構造上で直接組み立てられます。しかしヒト細胞では、γTuRCは細胞質内で予め組み立てられ、各種の組織化中心へと輸送されます。余分な微小管の形成を避けるため、ヒト版は正しい微小管の幾何学に合わない曲がった開いた形で保持され、大部分は不活性なままです。
アクセラレータのような補助因子
いくつかの細胞内タンパク質がγTuRCの活性を高めることができます。その多くはCM1と呼ばれる短い領域を共有し、これがリング複合体に直接結合します。個々の成長イベントを記録する高感度顕微鏡を用いて、著者らはガラス表面上で精製したヒトγTuRC分子が微小管を立ち上げようとする様子を観察しました。単独では複合体は鈍く、動きが遅かったのに対し、ヒトタンパク質由来のCM1断片を加えると核生成は劇的に加速し、通常のチューブリンを用いた場合で100倍以上、成長を促すよう設計されたチューブリン変異体を使うとさらに顕著でした。高濃度のCM1では、表面上のほぼすべてのγTuRCが最終的に作動し、この補助因子が集団全体を完全に活性化できることを示しました。
結合と管成長の相乗効果
CM1に蛍光タグを付けることで、研究者は個々のγTuRC分子にいつ結合したかと各微小管の成長がいつ始まったかを時間的に追跡できました。CM1結合と同時に微小管が現れることもありましたが、多くの場合は数分の遅延がありました。これはCM1の結合だけでは不十分で、複合体が形状をいくつか変化させる必要があることを示唆します。著者らは、CM1がγTuRCを「準備状態」にして最初のチューブリンサブユニットの組み立てを容易にすると提案しました。その後、管の成長自体が複合体を完全に閉じた対称的なリングへと駆動し、標準的な13本構造に一致させます。つまり、テンプレートと成長する管が協調してオフからオンへの切り替えを完了するのです。
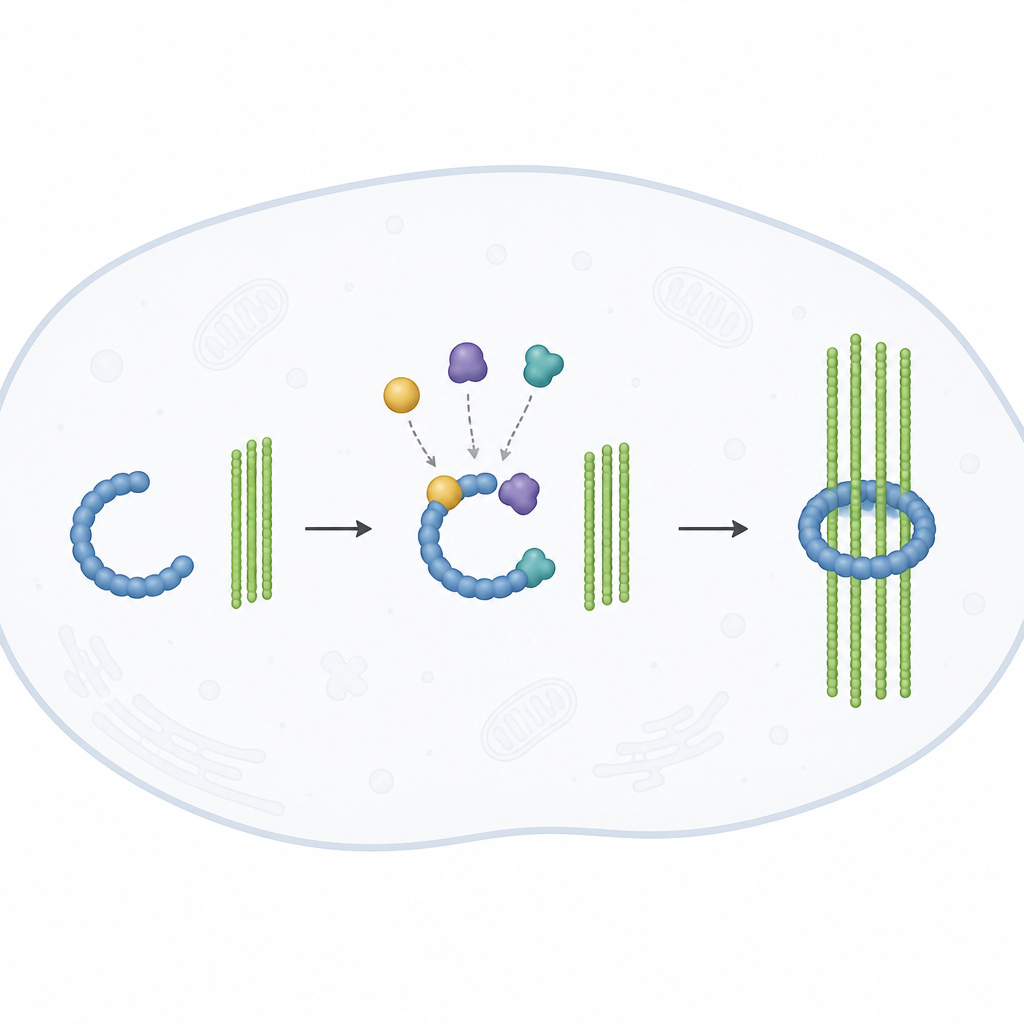
リングが閉じる瞬間のスナップショット
これらの形状変化を詳細に見るため、著者らはクライオ電子顕微鏡を用いました。この手法で、急速冷凍した分子を原子近傍の分解能でイメージングしました。彼らはCM1に結合し、かつ新たに形成された微小管の基部で既にキャップされているγTuRCを捉え、通常のチューブリンまたは成長に有利な変異体のいずれを使っても観察しました。いずれの場合も、リング複合体は完全に閉じ、その構成要素は標準的な13本の配列に一致する規則的なパターンで整列していました。これは少なくともヒト細胞では、効率的な核生成がリングの完全な閉鎖を伴うことを裏付けます。カエルを使った以前の研究は、脊椎動物のγTuRCが部分的に開いたままで特異な管形状を作る可能性を示唆していましたが、新しい研究はヒトの複合体が能動的な核生成時に完全に適合する姿をとると主張します。
リングをロックするラッチと内側の支え
高解像度で、著者らはリングを閉じるハードウェアのように機能する特定のタンパク質断片を同定できました。一つのγTuRCサブユニットから伸びる柔軟な伸長領域が小さなパートナータンパク質と協働して、ラッチと呼ばれる構造を形成します。このラッチは開いたらせんの後端から反対側に届き、リング内の最初のγ-チュブリンと新たに出現する微小管の最初のα-チュブリンの両方を把握します。同時に、CM1二量体が円錐外側の隣接サブユニット間に橋を形成し、特に一つの特別な部位で強い接触を作ります。そこから、余分なループが継ぎ目を越えて伸び、遠位側のγ-チュブリンに触れます。円錐内部では、内部の補強要素の一部であるアクチン分子が新しい位置に動き、閉鎖を妨げなくなって終端サブユニットと接触し、閉じたリングの安定化を助けます。
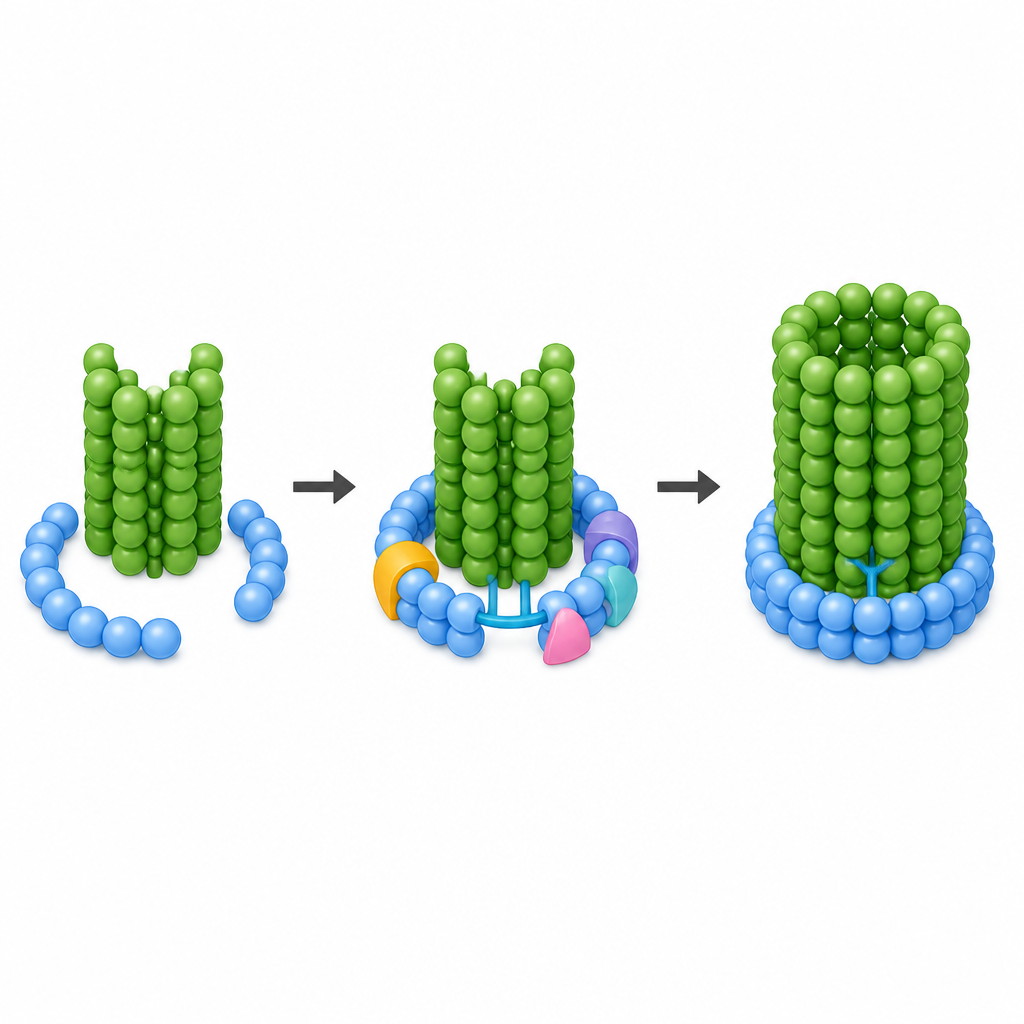
この分子スイッチが重要な理由
専門外の読者に向けた本研究のメッセージは、ヒト細胞が内部の管をいつどこで作るかを制御するために巧妙な安全機構を使っているという点です。γTuRC機械は安全な不活性形で組み立てられます。CM1と呼ばれる補助領域がそこにドッキングして緩めますが、完全な活性化は最初のチューブリンサブユニットが到着し、微視的なラッチと支えがリングを完全な円にロックしたときにのみ起こります。この連携した作用により、新しい微小管は正しい幾何学と正しい場所で始まり、正確な細胞分裂と細胞内部の秩序ある組織化を支えます。
引用: Serna, M., Brito, C., Speroni, S. et al. Structural basis of human γTuRC closure during CM1-activated microtubule nucleation. Nat Commun 17, 4488 (2026). https://doi.org/10.1038/s41467-026-70773-w
キーワード: 微小管核生成, γTuRC, CM1モチーフ, クライオ電子顕微鏡, 細胞分裂