Clear Sky Science · ja
細胞型特異的エンハンサーが自然免疫および獲得免疫のタイプ3リンパ球におけるIL-22発現を制御する
体の境界で守る者たち
私たちの皮膚や腸は常に微生物や刺激因子にさらされていますが、大半の場合それらは無傷で健康が保たれます。その重要な防御因子の一つがIL-22という分子で、バリア面近傍に存在する免疫細胞によって作られます。本研究は一見単純だが重要な問いを投げかけます:免疫細胞はいつ、どこで、どれだけのIL-22を作るかをどのように決めているのか?その答えは、感染からの防御を精密に調節しつつ、乾癬のような慢性炎症性疾患のリスクを抑えるDNA中の“ディマー(調光)スイッチ”を明らかにします。
なぜIL-22が日常の健康に重要か
IL-22は免疫系からバリア組織(腸、肺、肝、皮膚、乳房など)への直接のホットラインのように働きます。IL-22が上皮細胞の受容体に結合すると、抗菌防御の強化、組織修復の促進、細胞の損傷からの生存促進をもたらすプログラムが誘導されます。例えば腸では、IL-22はマウスモデルで有害な大腸菌株に相当するCitrobacter rodentiumなどの細菌を撃退するのに不可欠です。IL-22が少なすぎると組織は感染や損傷に脆弱になり、多すぎると慢性炎症やがんを助長する可能性があります。痛みを伴う皮膚疾患である乾癬の患者では、IL-22が高値を示し病勢と相関することが多いです。IL-22やそのシグナル経路を標的とする薬剤が臨床試験段階にあるため、IL-22がDNAレベルでどのように制御されるかを理解することは基礎的・医療的に重要です。
単一の重要分子のための隠れたDNAスイッチ
著者らは「タイプ3リンパ球」として知られる免疫細胞群、すなわち自然リンパ球(ILC3)や関連するヘルパーT細胞(Th17/Th22)に着目しました。これらの細胞は類似した役割を共有しますが、応答の速さや活性化の契機は異なります。モデルILC3細胞株で強力なCRISPR干渉スクリーニングを用い、IL-22遺伝子周辺の約150万塩基にわたる領域を走査しました。免疫細胞で活性がありそうに見える何百ものDNA領域を系統的にサイレンシングし、どれがIL-22産生に本当に必要かを問いかけました。多くの候補の中で、特に重要な二つのエンハンサー—短い制御DNA断片—が浮かび上がりました。E22-1とE22-2と名付けられたこれらはIL-22遺伝子の上流に数万塩基離れて位置し、マウスとヒトで保存されていることから進化的に重要な役割が示唆されます。
自然免疫と獲得免疫の間での役割分担
これらのエンハンサーが生体内でどのように機能するかを調べるため、研究者らはE22-1、E22-2、あるいは両方を欠くマウスを作製しました。驚くべきことに、両方のスイッチを欠失させると、ILC3だけでなく特定のT細胞やナチュラルキラー様細胞など他のIL-22産生細胞でもIL-22産生が大幅に低下し、近傍の遺伝子は損なわれませんでした。これらのマウスは腸上皮における常時の抗菌遺伝子の発現が低く、腸感染に対して脆弱でした:C. rodentiumで挑戦した後、通常のマウスより体重減少が大きく、菌の負荷も高かったのです。E22-1単独の除去では、IL-22は自然免疫・獲得免疫双方のタイプ3細胞で広く低下し、事実上二重ノックアウトと同様の表現型を示しました。対照的にE22-2の欠失は主にILC3と一部のγδ T細胞におけるIL-22を損ない、従来のヘルパーT細胞にはほとんど影響を与えませんでした。E22-2だけが蛍光マーカーを駆動するレポーターマウスではこのパターンが確認されました:多くの組織でILC3に強い活性、他のリンパ球では弱または希な活性、古典的ヘルパーT細胞では感染時でもほとんど活性が見られませんでした。
保護から乾癬へ
IL-22が過剰だと有害になり得るため、研究班はこれらのエンハンサーが皮膚炎にどう影響するかを次に調べました。彼らはイミキモドという免疫刺激剤を反復塗布すると乾癬様病変を誘導する標準的なマウスモデルを用いました。正常マウスは厚く赤く鱗状の耳を発症し皮膚で高いIL-22レベルを示しました。E22-1、E22-2、あるいは両方を欠く三種のエンハンサー欠損株はいずれも保護されており:耳は薄く、発赤や鱗屑は軽度で、皮膚のIL-22発現は大幅に低下していました。一方で関連サイトカインのIL-17AやIL-17Fのレベルはほぼ同等でした。特に“自然免疫寄り”のエンハンサーであるE22-2の欠失だけで、広く作用するE22-1の欠失と同等の保護効果が得られたことは、ILC3やγδ T細胞など皮膚常在の自然免疫細胞由来のIL-22だけで乾癬様疾患を駆動するのに十分であることを示唆します。
スイッチが分子レベルでどのように働くか
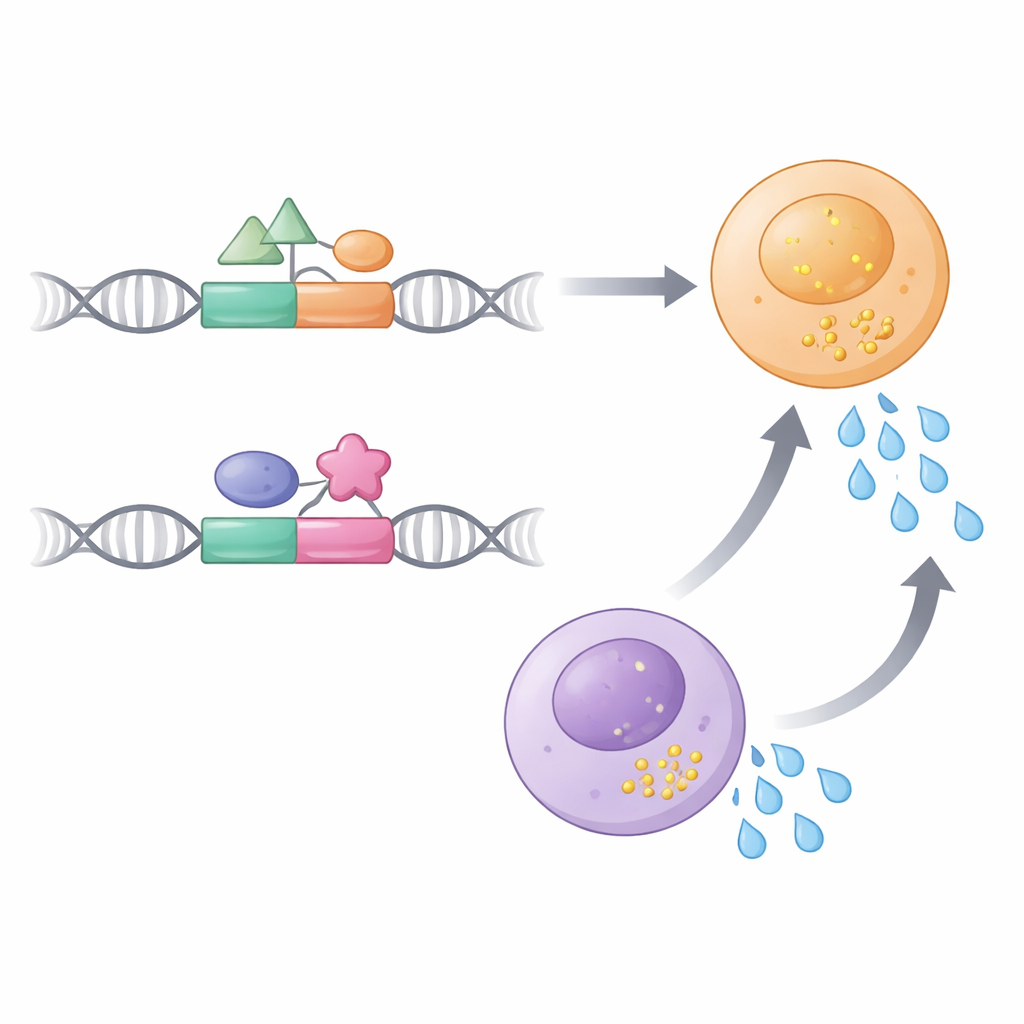
さらに踏み込んで著者らはこれらのエンハンサーの内部構造を解剖しました。E22-1とE22-2の両方には転写因子の結合部位が含まれており、転写因子はシグナルを感知して遺伝子をオンにします。ILC3では、E22-2は自然リンパ系の分化を導くことで知られるRunx3の複数の結合部位と、AP-1ファミリーの協働因子に強く依存していました。クロモソーム折りたたみ実験は、E22-2が機能するにはIL-22プロモーターへの物理的接触が必要であり、この接触自体がE22-1の存在に依存していることを示しました。E22-1を除去するとE22-2のアクセス可能性と遺伝子へのループ形成能力が著しく低下し、「依存的エンハンサー(contingent enhancer)」の関係が明らかになりました:E22-1はE22-2が完全に機能するための地盤を整えます。重要なひねりは、E22-1とは異なりE22-2はILC3やTh17/Th22細胞で強く発現する転写因子RORγtの有効な結合部位を欠いている点です。著者らが実験的にE22-1の強いRORγt結合部位をE22-2に差し替えると、E22-2はヘルパーT細胞で突然強固な活性を獲得しました。これは単一の微細なDNAモチーフがエンハンサーの活性を自然免疫細胞に限定し、獲得免疫側を除外することがあり得ることを示唆します。
防御と損傷のバランス
総じて本研究は、二つの協働するDNAエンハンサーが異なる種類のリンパ球におけるIL-22出力を細かく調節する層構造の制御系を明らかにしました。E22-1は自然免疫および獲得免疫の双方に対する広く共有されたスイッチとして働き、E22-2はILC3と一部のγδ T細胞に調整されたスイッチとしてバリア面での基礎的防御とトラブル時の迅速な自然免疫ブーストを提供します。これらの共同作用は腸の抗菌防御を維持し、過剰に活性化されると炎症性皮膚疾患に寄与します。この制御回路を詳細に地図化することで、免疫系が同一分子IL-22を用いて組織を守る一方で、誤調節時には慢性炎症を促進し得る仕組みが説明されました—これはIL-22を全体的に抑えるのではなく、そのDNA制御スイッチのレベルで調節するより精密な治療法を導く手掛かりとなるでしょう。
引用: Saini, A., Hopkins, L.S., Serna, V.A. et al. Cell type-specific enhancers regulate IL-22 expression in innate and adaptive type 3 lymphoid cells. Nat Commun 17, 3960 (2026). https://doi.org/10.1038/s41467-026-70636-4
キーワード: IL-22, 自然リンパ球, 遺伝子エンハンサー, バリア免疫, 乾癬