Clear Sky Science · he
דִגּוּם מתמטי של קוטביות מישורית של תאים: עקרונות, גישות ושאלות פתוחות
כיצד תאים מסתדרים על כיוון משותף
איברים רבים בגופנו, מהאוזן הפנימית ועד דרכי הנשימה והכליות, מתפקדים בזכות שכבות תאים שכולן "פונות" באותו הכיוון על פני שטח הרקמה. יישור מתואם זה, הנקרא קוטביות מישורית של תאים, משמש כקומפאס פנימי שמנחה כיצד רקמות גדלות, מתעוותות ומתפקדות. כשקומפאס זה משתבש, התוצאות עלולות להיות חמורות ולתרום למומים מולדים, לאובדן שמיעה, לבעיות נשימה ולעיוותי עמוד שדרה. מאמר זה סוקר כיצד מדענים משתמשים במודלים מתמטיים ומחושבים כדי להבין כיצד מולקולות בודדות בתוך תאים משתפות פעולה ליצירת סדר בקנה מידה גדול כזה.
מדוע הכיוון ברמת הרקמה חשוב
קוטביות מישורית של תאים (PCP) מתארת כיצד תאים בדף דק מסתדרים בכיוון משותף, לצדדי הרקמה ולא קדימה־אחורה. בעכברי פרי (זבוב הפרי) משכה ה‑PCP תשומת לב כאשר שערות זעירות על הכנף פנו כולן באותו הכיוון. דפוסי כיוון דומים מופיעים גם בוורטברטים: רקמות עצב סוגרות נכון רק אם תאים נמתחים ונכנסים לסידור מיושר; תאי שערה באוזן הפנימית חייבים להיות מונחים בדיוק כדי לקלוט קול; ריסים בדרכי הנשימה וחדרי המוח חייבים להכות בסנכרון להזזת נוזלים; וזקיקי שיער בעור נוטים לדפוסים מתואמים. פגמים בגנים שמווסתים PCP קשורים לפגמים בצילינדר העיצבי (neural tube defects), להפרעות של השלד כמו תסמונת רובינוב, לעיוותי כליה ולקיפולי עמוד שדרה. כיוון ש‑PCP מעצבת איברים הן בעובר והן בבוגר, החוקרים מבקשים להבין כיצד מולקולות בתוך כל תא משלבות מידע מקומי עם רמזים מרחיקי טווח כדי לשמור על יישור תקין של הרקמות.
שתי מערכות קומפאס מולקולריות
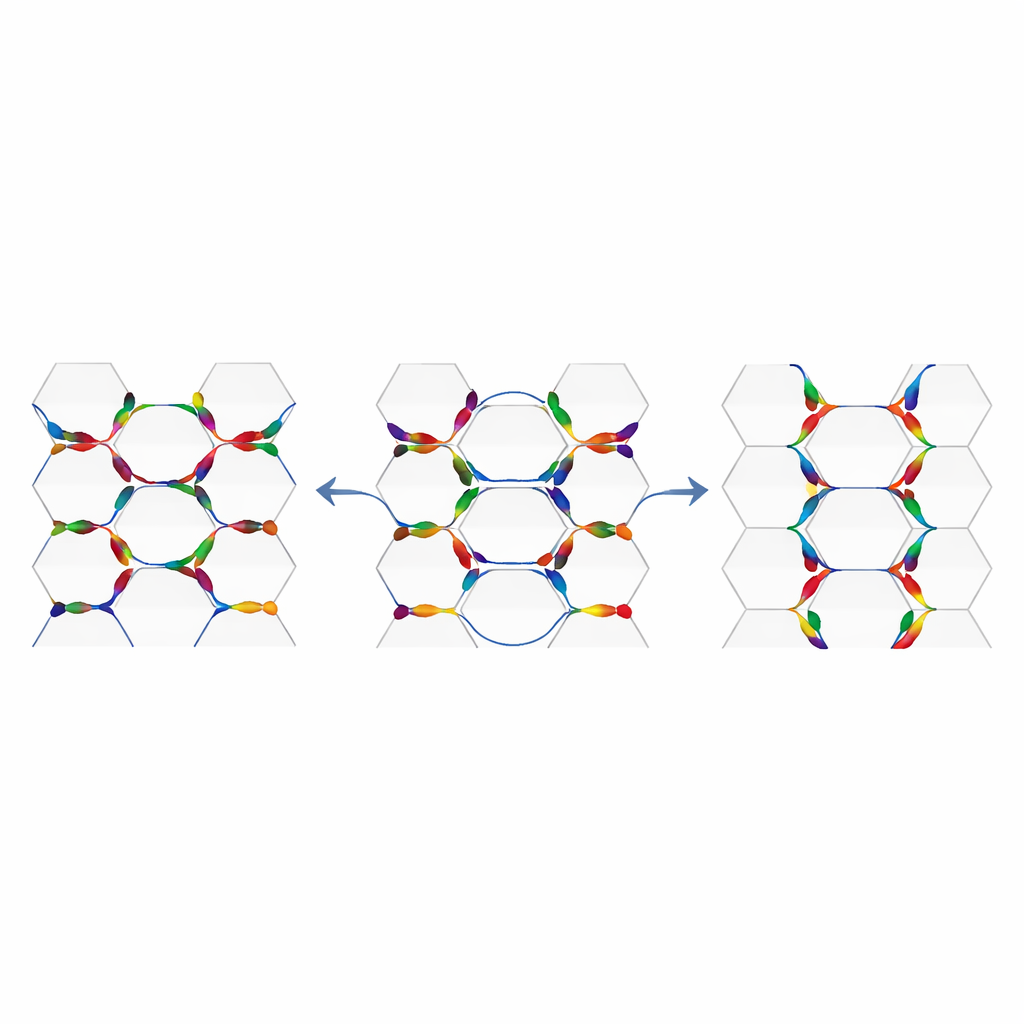
ביולוגים זיהו שני מערכי חלבונים משולבים שפועלים יחד לבניית PCP. האחד, הידוע כמודול הליבה, כולל חלבונים ממברנליים וציטופלזמטיים שמצטברים באופן אסימטרי על צדדים מנוגדים של כל תא. הצבירות האלה בין תאים שכנים מתקשרות בחצאי‑ממברנה ומאפשרות לתאים להשוות את כיוונם עם השכנים. מודול שני, גלובלי יותר, מורכב מחלבוני קדחרין גדולים הנקראים Fat ו‑Dachsous, בתוספת אנזים משנות בשם Four‑jointed. רכיבים אלה מבוטאים במדרונים ברמת רקמה, כך שצד מסוים של הרקמה רואה מעט יותר מאחד החלבונים מהצד השני. אותה דרגתיות משפיעה בעדינות על המקום בו מתחבאים קומפלקסים בכל תא, ועוזרת ליישר את המנגנון המקומי עם ציר האיבר הכולל. כיצד בדיוק שני המודולים האלה מתקשרים — האם אחד מזין את השני ברצף או פועלים במקביל — נותר תחום מחקר פעיל.
עדשות מודלים שונות לאותה בעיה
הסקירה מסבירה כיצד נבנו מספר משפחות של מודלים מתמטיים, לכל אחת יתרונות משלה, לחקר PCP. מודלי Potts תאיים מייצגים רקמות על רשת, ועוקבים אחר צורות תאים מפורטות ומיקומי חלבוני הקוטביות לאורך קצוות התאים; הם מראים כיצד רמזים זמניים או מקומיים יכולים להגבר לסדר בטווח ארוך. מודלי רשת פטרי מתמקדים באירועי קשירה וניתוק בדידים של חלבונים באזורים ממברנליים קטנים, ותופסים את האופי הסטוכסטי של מפגשי מולקולות. מודלים מבוססי סוכנים מטפלים בכל תא כישות שהרמות והאינטראקציות של חלבוני הממברנה שלה משתנות בזמן לפי חוקים דמויי‑תגובה; מודלים אלה משמשים לרוב לחקור כיצד משוב, רעש, מדרונים וגיאומטריית תאים משתלבים ליצירת דפוסי רקמה. מודלים פננומנולוגיים דוחסים את כל הפרטים המולקולריים לווקטורים פשוטים של קוטביות ולפונקציות דמויות‑אנרגיה, תוך הלוואת רעיונות ממגנטיות כדי לנתח מתי מופיעים או מתפרקים דפוסים מסודרים. לבסוף, תיאוריות רציפות מעגלות את הפינות על פני תאים בודדים כדי לתאר קוטביות כשדה רציף הציית למשוואות דיפרנציאליות, ומאפשרות תובנות אנליטיות על רקמות גדולות וקשרים למערכות יצירת דפוסים אחרות.
מה המודלים מלמדים אותנו
בין הגישות האלה עולות כמה מסקנות כלליות. משוב מקומי בין צמתים שכנים יכול ליצור אסימטריה אפילו מרמזים גלובליים חלשים, אך מדרונים של חלבונים כגון Fat ו‑Dachsous עוזרים ליישר את הקוטביות על פני דומיינים גדולים ולמצב דפוסים עמידים יותר לתנודות אקראיות. תהליכים פנימיים לתא, כמו תנועת חלבונים ואינטראקציות בתוכו, הופכים חשובים במיוחד כאשר האותות החיצוניים רעשיים או חלשים. צורת התא וגיאומטריית הרקמה גם הן חשובות: כאשר תאים מוארכים או רקמות נמתחות, הקוטביות נוטה להתיישר בכיוונים צפויים יחסית לעיוות זה, מרמזת על קשרים עמוקים בין מכניקה לאותות מולקולריים. מודלים מראים כיצד רעש מולקולרי אקראי או אי‑סדירות קבועות ברמות חלבון יכולים ליצור דפוסי קוטביות מסתחררים או אנטומיים, וכיצד שיבוטים של תאים מוטנטים יכולים להפריע לכיוון השכנים — התנהגות הידועה כלא‑אוטונומיה מרוחקת (domineering non‑autonomy) הנצפית בניסויים. במקביל, הנחות מזעריות שונות לגבי הפרטים המיקרוסקופיים מסוגלות להוליד התנהגות דומה בקנה מידה גדול, מה שמדגיש כמה קשה להסיק את כללי המולקולות המדויקים מתוך תצפיות ברמת הרקמה בלבד.
חידות פתוחות וכיווני מחקר עתידיים
למרות התקדמות משמעותית, הדגומים מדגישים כמה שאלות לא פתורות. האם שני המודולים של PCP פועלים בדרך כלל ברצף או במקביל, והאם התשובה משתנה מרקמה לרקמה? האם עדיף לתאר PCP כתהליך בשיווי משקל, כמו מגנטים שמסתדרים בשדה, או כמערכת מופעלת שאינה בשיווי משקל ודומה להסתדרות עופות? עד כמה זרימות רקמה, כוחות מכאניים וצורת האיבר משפיעים בחזרה על הקוטביות המולקולרית, והאם אפשר לקשר בין האלמנטים האלה במודלים מאוחדים חוצי‑קנה מידה? המחברים טוענים שהתשובה לשאלות אלה תצריך שיתוף פעולה הדוק בין תאוריה וניסוי, מונחה על‑ידי מודלים שמציעים ניבויים חדים הניתנים למדידה.
מה המשמעות לכך בבריאות ומחלה
ללא‑מומחים, המסר המרכזי הוא שמבנה מאורגן של האיברים שלנו נשען על אינספור החלטות מיקרוסקופיות שהחלבונים בגבולות התאים מקבלים, ומתמטיקה הופכת לכלי חיוני לפיצוח כיצד ההחלטות הללו מצטברות. בהשוואת מסגרות דִגּוּם שונות והתובנות שהן מספקות, סקירה זו מציירת מפת דרכים לבניית תורות חיזוי של קוטביות מישורית של תאים. תיאוריות כאלה עשויות בסופו של דבר לסייע להסביר מדוע מתרחשים מומים מסוימים, מדוע חלק מהרקמות עמידות יותר לנזק מאחרות, וכיצד אולי נוכל בעתיד לכוון את ארגון הרקמה ברפואה שיקומית או בטיפול במחלות.
ציטוט: Rizvi, M.S., Jolly, M.K. Mathematical modeling of planar cell polarity: principles, approaches, and open questions. npj Syst Biol Appl 12, 56 (2026). https://doi.org/10.1038/s41540-026-00679-2
מילות מפתח: קוטביות מישורית של תאים, רקמת אפיתל, דִגּוּם מתמטי, מורפוגנזה של רקמות, אותות תאיים