Clear Sky Science · fr
Commutation dépendante du nucléotide et reconnaissance de l'effecteur RIPb du facteur de susceptibilité de l'orge RACB
Comment un champignon détourne les défenses de la plante
Le mildiou poudreux est une maladie fongique courante qui recouvre les feuilles d'orge d'un duvet blanc et réduit les rendements. Cette étude explore en profondeur les cellules d'orge pour comprendre comment un petit interrupteur moléculaire, nommé RACB, peut être détourné afin que le champignon trouve plus facile de traverser la paroi externe de la plante. En révélant les formes et les mouvements exacts de cet interrupteur et de sa protéine auxiliaire RIPb, les chercheurs montrent comment un système de contrôle naturel de la plante est réutilisé pour favoriser l'infection.
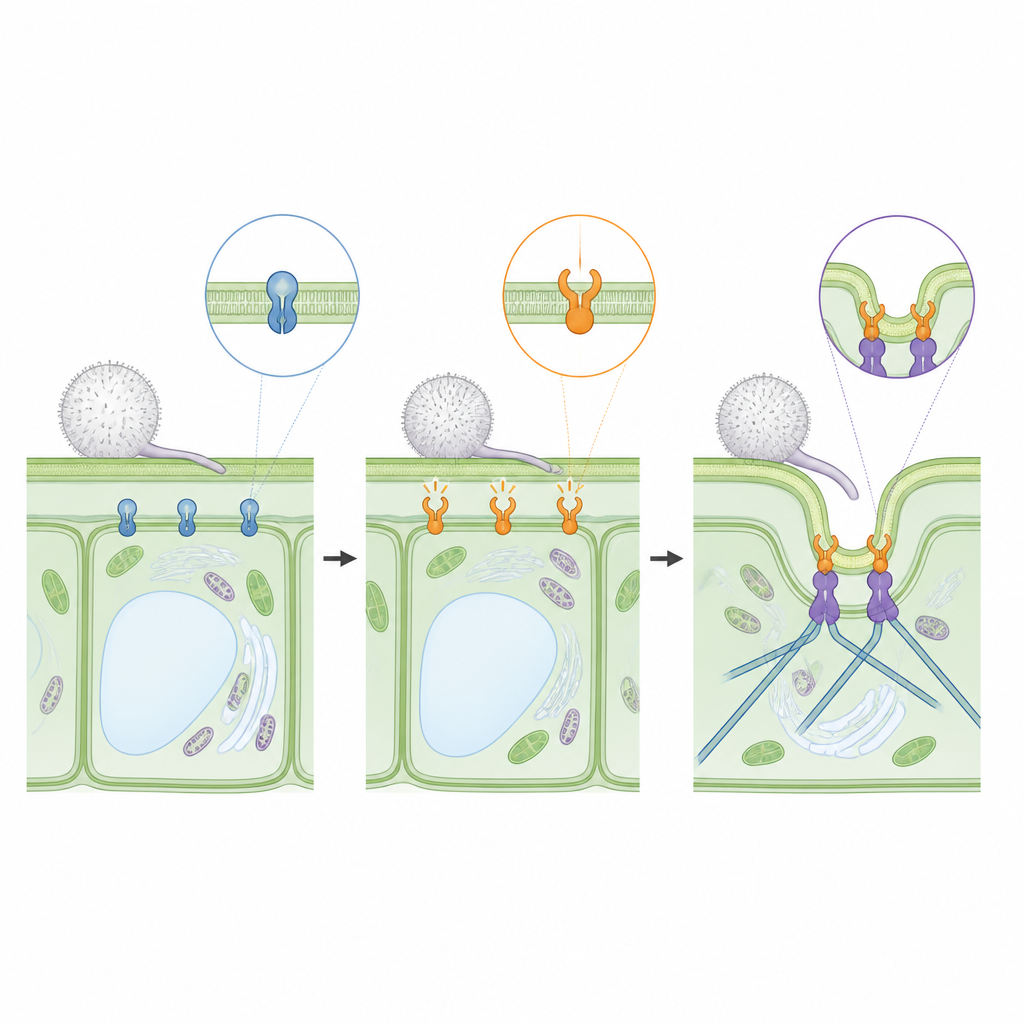
Un interrupteur moléculaire à la surface cellulaire
De nombreuses cellules, chez l'humain comme chez les plantes, s'appuient sur de petites protéines qui fonctionnent comme des interrupteurs marche/arrêt. Ces interrupteurs basculent entre une forme inactive et une forme active selon la petite molécule qu'ils portent. Chez l'orge, RACB est un de ces interrupteurs positionné à la face interne de la membrane cellulaire. Lorsqu'il est éteint, la plante est moins accueillante pour les maladies. Lorsqu'il est activé, la cellule réorganise sa structure interne, ce qui, chez l'orge, est associé à une plus grande susceptibilité au champignon du mildiou poudreux Blumeria hordei. Des travaux antérieurs avaient montré que le maintien permanent de RACB en position activée facilite l'entrée du champignon dans les cellules d'orge, tandis que sa répression rend l'infection plus difficile.
Capturer RACB en mouvement
L'équipe a utilisé une combinaison d'outils structuraux puissants pour observer RACB au détail atomique dans différents états. La cristallographie aux rayons X a fourni des instantanés de RACB lié à une molécule « éteinte » et à deux molécules « pseudo-activées » qui imitent sa forme active. La résonance magnétique nucléaire et une technique suivant l'échange des hydrogènes avec l'eau lourde ont ensuite révélé la flexibilité de différentes parties de la protéine en solution. Ensemble, ces expériences ont montré que deux régions clés de RACB, appelées switch I et switch II, déplacent progressivement leur position et leur flexibilité à mesure que la protéine passe de l'état éteint à partiellement activé puis totalement activé. Plutôt que de se comporter comme un simple interrupteur binaire, RACB explore une gamme de conformations, les formes actives présentant des mouvements internes plus forts et plus rapides, surtout autour des régions switch.
Comment RACB saisit sa protéine auxiliaire
RACB n'agit pas seul. Lors de l'attaque fongique, il recrute une protéine d'orge appelée RIPb qui peut se lier à la fois à la membrane et aux fibres de soutien internes appelées microtubules. À l'aide de mesures d'affinité et de travaux structuraux supplémentaires, les chercheurs ont montré que RIPb reconnaît uniquement la forme active, entièrement commutée, de RACB. Ils ont identifié un court segment dans RIPb, de séquence QWRKAA, qui se niche entre les deux régions switch de RACB. Dans des structures cristallographiques à haute résolution, ce court segment de RIPb forme une hélice dont les chaînes latérales établissent des contacts serrés avec RACB, verrouillant les switchs dans leur arrangement actif. Lorsque l'équipe a modifié deux positions critiques dans ce motif, RIPb n'a plus pu se lier à RACB ni en éprouvettes, ni dans des cellules de levure, ni dans des cellules d'orge vivantes, et les signaux fluorescents rapportant leur interaction ont quasiment disparu.
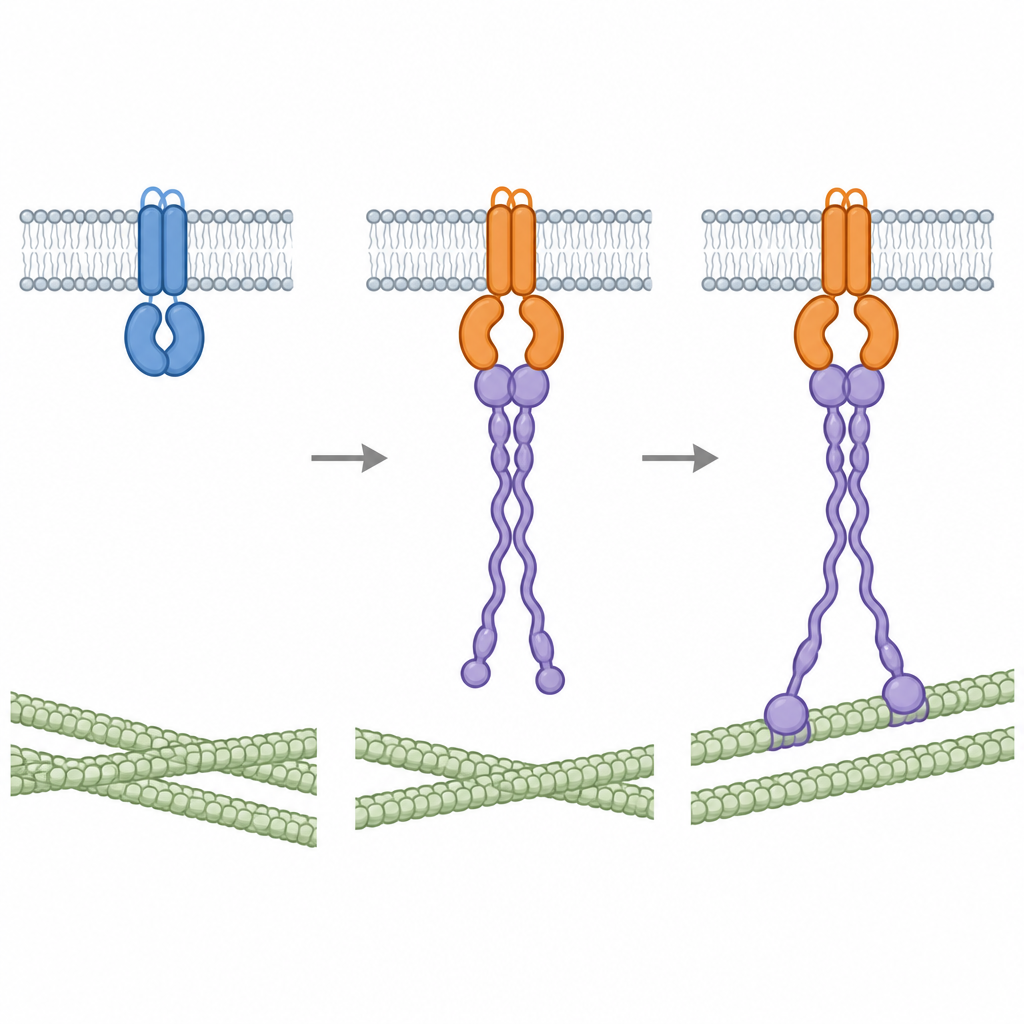
Construire un pont entre la membrane et l'armature interne
En combinant leurs structures avec des simulations informatiques, les auteurs ont élaboré un modèle de la façon dont RACB et RIPb se positionnent ensemble sur la face interne d'une cellule d'orge. RACB est ancré à la membrane par une queue lipidique et une zone de charges positives, tandis que RIPb forme une tige dimérique qui porte également des charges positives près de son extrémité. Dans le modèle, une paire de molécules RACB retient une paire de molécules RIPb, avec leurs queues enfouies dans la membrane et les extrémités distales de RIPb s'étendant en direction des microtubules à l'intérieur de la cellule. Cette organisation fournit un pont physique qui pourrait aider à remodeler la membrane et à guider l'ossature interne précisément à l'endroit où le champignon tente d'envahir.
Ce que cela signifie pour la protection des cultures
L'étude conclut que l'interrupteur RACB de l'orge est gouverné par des changements subtils de conformation et de dynamique, et que le champignon tire avantage du stabilisation de RACB dans sa forme pleinement active par RIPb. Le segment conservé QWRKAA de RIPb agit comme une clé qui s'insère dans la serrure RACB active, reliant la membrane cellulaire à l'armature interne nécessaire au remodelage local. Pour les non-spécialistes, cela signifie que le champignon ne force pas simplement la porte, mais exploite astucieusement les mécanismes de contrôle de la plante pour l'ouvrir. Comprendre ce mécanisme détaillé suggère des voies futures pour sélectionner ou concevoir des plants d'orge chez lesquels cette interaction est affaiblie, de sorte que le même interrupteur moléculaire soutienne la croissance et la défense normale sans fournir au champignon une porte d'entrée facilitée.
Citation: Mohamadi, M., Bradai, M., Janowski, R. et al. Nucleotide-dependent switching and RIPb effector recognition of the barley susceptibility factor RACB. Commun Biol 9, 691 (2026). https://doi.org/10.1038/s42003-026-10316-7
Mots-clés: immunité de l'orge, mildiou poudreux, petite GTPase, cytosquelette, interaction plante-pathogène