Clear Sky Science · fr
β-arrestine1 orchestre la signalisation endosomale pour réguler le contrôle traductionnel de l’alignement circadien par la lumière
Comment la lumière garde nos horloges internes à l’heure
Tous ceux qui ont souffert du décalage horaire ou de soirées tardives devant un écran savent ce qui arrive quand l’horloge corporelle se désynchronise du jour. Cette étude examine comment un assistant moléculaire spécifique à l’intérieur des cellules cérébrales, appelé β-arrestine1, traduit la lumière entrant par les yeux en ajustements précis de l’horloge maîtresse du cerveau, maintenant les rythmes quotidiens alignés sur le lever et le coucher du soleil.
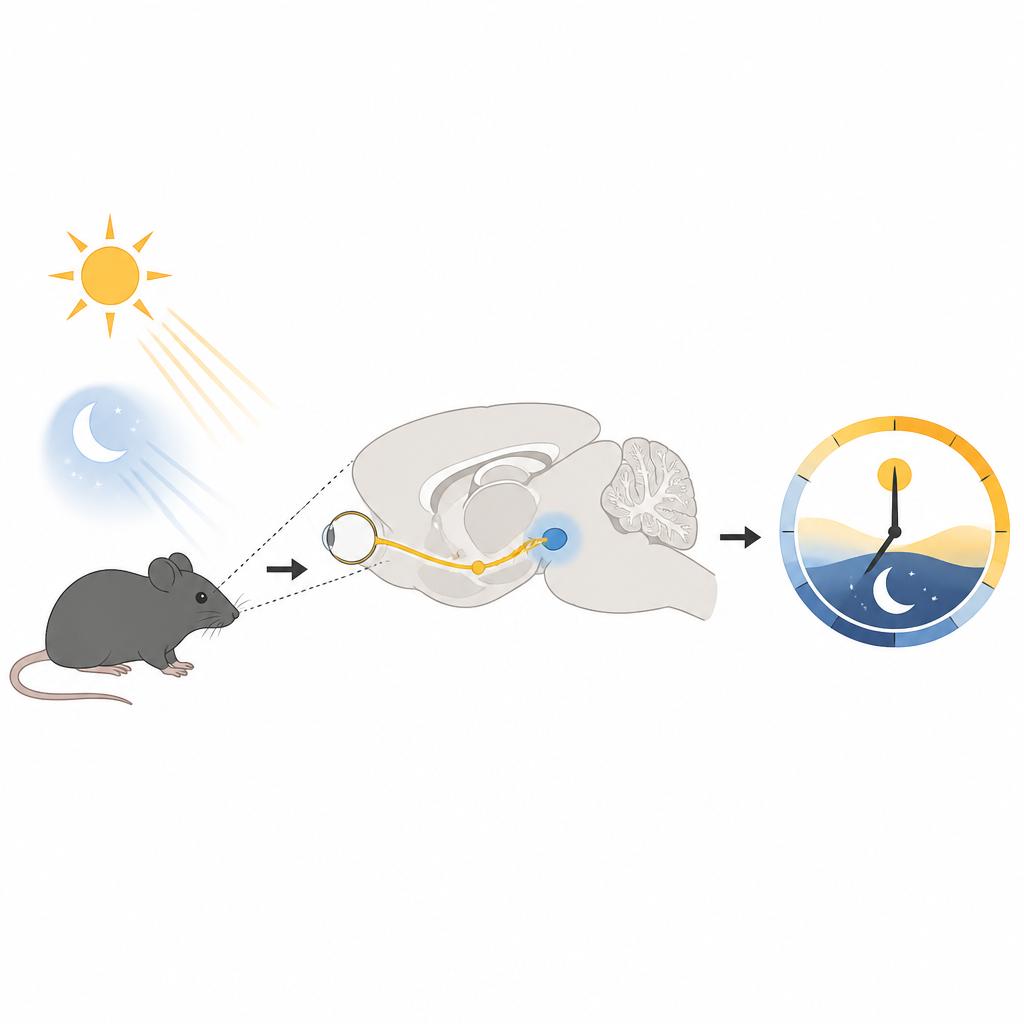
Présentation du gardien du temps cérébral
Chez les mammifères, une minuscule région située au fond du cerveau, le noyau suprachiasmatique, joue le rôle d’horloge maîtresse. Il coordonne les cycles quotidiens de sommeil, de sécrétion hormonale et de température corporelle. Cette horloge est réinitialisée par des signaux lumineux qui voyagent de l’œil le long d’une voie spécialisée jusqu’à cette région. À l’intérieur de ces cellules horlogères, un ensemble de gènes et de protéines monte et descend en boucle de 24 heures, et la lumière peut avancer ou retarder cette boucle pour que notre heure interne corresponde au monde extérieur.
Un assistant moléculaire avec une mission particulière
Nombre des signaux induits par la lumière dans l’horloge utilisent des récepteurs de surface appartenant à une vaste famille connue sous le nom de récepteurs couplés aux protéines G. L’un d’eux, appelé PAC1, répond à un messager libéré par des fibres rétiniennes quand la lumière frappe l’œil. Les chercheurs se sont concentrés sur deux protéines auxiliaires étroitement apparentées, la β-arrestine1 et la β-arrestine2, connues pour réguler l’activation, l’inactivation et le déplacement de ces récepteurs à l’intérieur des cellules. En étudiant des souris dépourvues de l’une ou l’autre, ils ont constaté que seule la β-arrestine1 était cruciale pour des réponses normales à la lumière, notamment la vitesse d’adaptation à un décalage horaire simulé et l’amplitude du déplacement des rythmes d’activité après une courte impulsion lumineuse nocturne.
Les signaux lumineux se déplacent à l’intérieur de la cellule
L’équipe a découvert que la β-arrestine1 fait plus que simplement éteindre les récepteurs de surface. Chez des souris normales, un éclair lumineux nocturne provoquait l’internalisation des récepteurs PAC1 des neurones horlogers dans de petites vésicules internes appelées endosomes. Ces endosomes servent de centres de signalisation, où la β-arrestine1 aide à assembler une chaîne d’interrupteurs protéiques, notamment une voie impliquant ERK, RSK1 et une protéine ribosomale appelée S6. Cette cascade stimule la machinerie de synthèse protéique de la cellule au moment opportun. Chez les souris dépourvues de β-arrestine1, les récepteurs PAC1 ne parvenaient pas à se localiser efficacement dans les endosomes et l’activation de cette voie de signalisation interne était fortement réduite.
Des signaux aux nouvelles protéines horlogères
La réinitialisation de l’horloge nécessite non seulement l’activation des gènes mais aussi la production suffisante de leurs produits protéiques. Les auteurs ont montré que, bien que la lumière déclenche encore des poussées normales d’activité génique chez les souris déficientes en β-arrestine1, la production effective des protéines clés de l’horloge, PER1 et PER2, était atténuée dans le noyau de l’horloge maîtresse. En utilisant une méthode qui marque les protéines nouvellement synthétisées, ils ont constaté que la lumière augmente normalement la synthèse protéique globale dans la région horlogère, mais que cette augmentation disparaît en l’absence de β-arrestine1. Cela souligne un rôle spécifique de la β-arrestine1 dans le contrôle de la traduction, l’étape où la machinerie de synthèse lit les messages génétiques et assemble de nouvelles protéines.
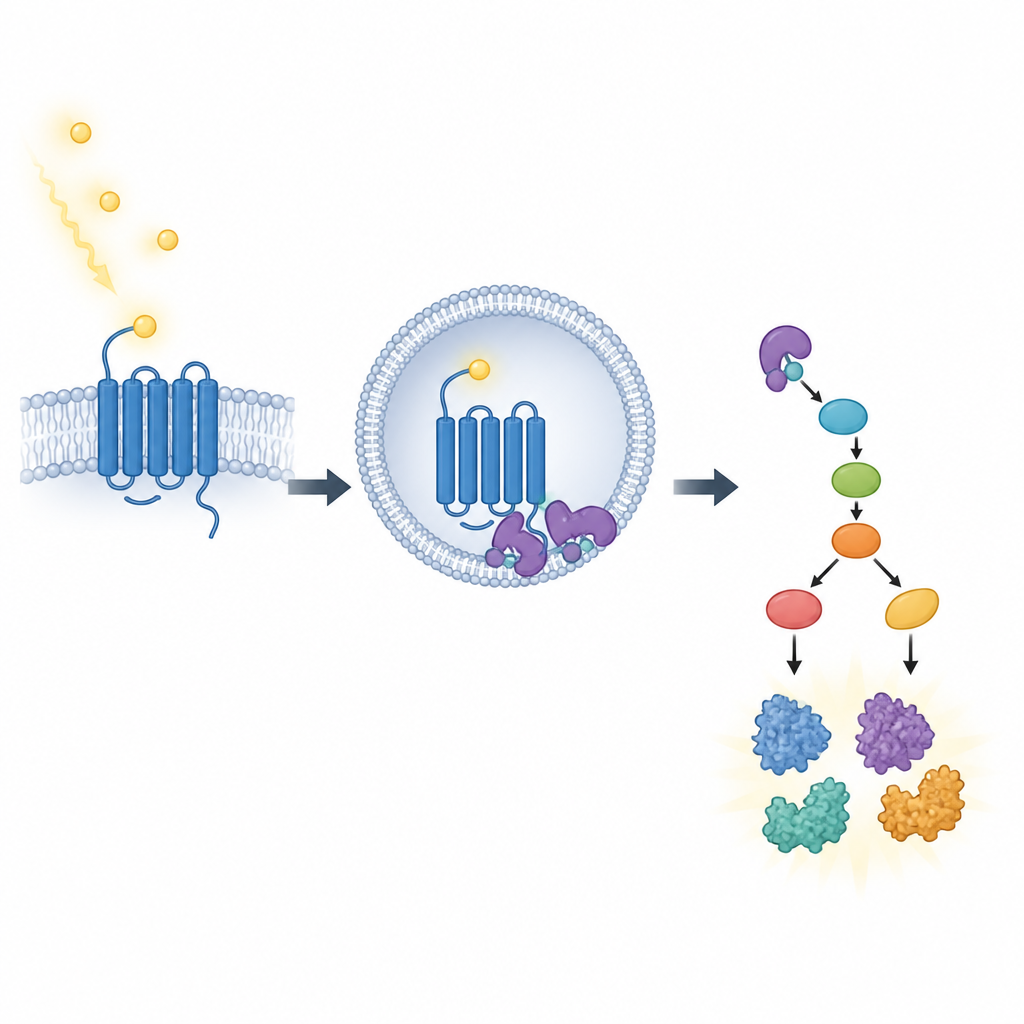
Équilibrer signaux de surface et signaux internes
L’étude a également évalué la contribution des voies de signalisation plus traditionnelles au niveau de la surface cellulaire par rapport à ces signaux endosomaux internes. En utilisant des médicaments pour bloquer différentes branches de la voie dans des coupes cérébrales et des cultures cellulaires, les chercheurs ont trouvé que la signalisation issue des endosomes apportait la contribution la plus importante à l’activation de la voie ERK en réponse à des stimuli imitant la lumière. Les signaux restés à la surface cellulaire par d’autres voies jouaient des rôles plus modestes et de soutien. En l’absence de β-arrestine1, certaines réponses basées à la surface persistaient, ce qui explique pourquoi l’activité génique précoce était préservée même si la production protéique était compromise.
Pourquoi cela compte pour la vie quotidienne
Dans l’ensemble, ces résultats révèlent que les endosomes à l’intérieur des cellules horlogères agissent comme d’importantes stations relais pour l’information lumineuse, et que la β-arrestine1 en est une coordinatrice clé. Plutôt que de simplement éteindre les récepteurs, la β-arrestine1 les oriente vers l’intérieur pour déclencher un programme de synthèse protéique qui permet à l’horloge de se réinitialiser correctement. Pour un lecteur non spécialiste, cela signifie que notre capacité à nous adapter à de nouveaux fuseaux horaires ou à des rythmes d’éclairage irréguliers dépend non seulement du fait que notre cerveau perçoive la lumière, mais aussi de la manière dont cette lumière active la machinerie interne de la cellule pour fabriquer les bonnes protéines horlogères au bon moment.
Citation: Mascarenhas, B., Seecharran, S., Boehler, N.A. et al. β-arrestin1 orchestrates endosomal signaling to regulate translational control of circadian light entrainment. Commun Biol 9, 645 (2026). https://doi.org/10.1038/s42003-026-09905-3
Mots-clés: rythmes circadiens, β-arrestine1, noyau suprachiasmatique, alignement par la lumière, récepteur PAC1