Clear Sky Science · fr
La condensation des paraspeckles est contrôlée par la polymérisation de TDP-43 et liée à la neuroprotection
Comment de minuscules gouttelettes nucléaires peuvent protéger les cellules cérébrales
À l’intérieur de chacune de nos cellules, en particulier des neurones, se trouvent de minuscules gouttelettes de type liquide appelées paraspeckles. Ces structures se forment autour d’une longue molécule d’ARN nommée NEAT1_2 et aident les cellules à faire face au stress. Dans de nombreuses maladies neurodégénératives, une autre protéine, TDP-43, se dérègle et s’agrège, mais son rôle quotidien dans les noyaux sains restait énigmatique. Cette étude révèle comment TDP-43 contrôle physiquement l’apparition et le comportement des paraspeckles, et comment de petites modifications de ce système de contrôle peuvent faire basculer l’équilibre entre la survie neuronale et la dégénérescence dans des troubles comme la sclérose latérale amyotrophique (SLA). 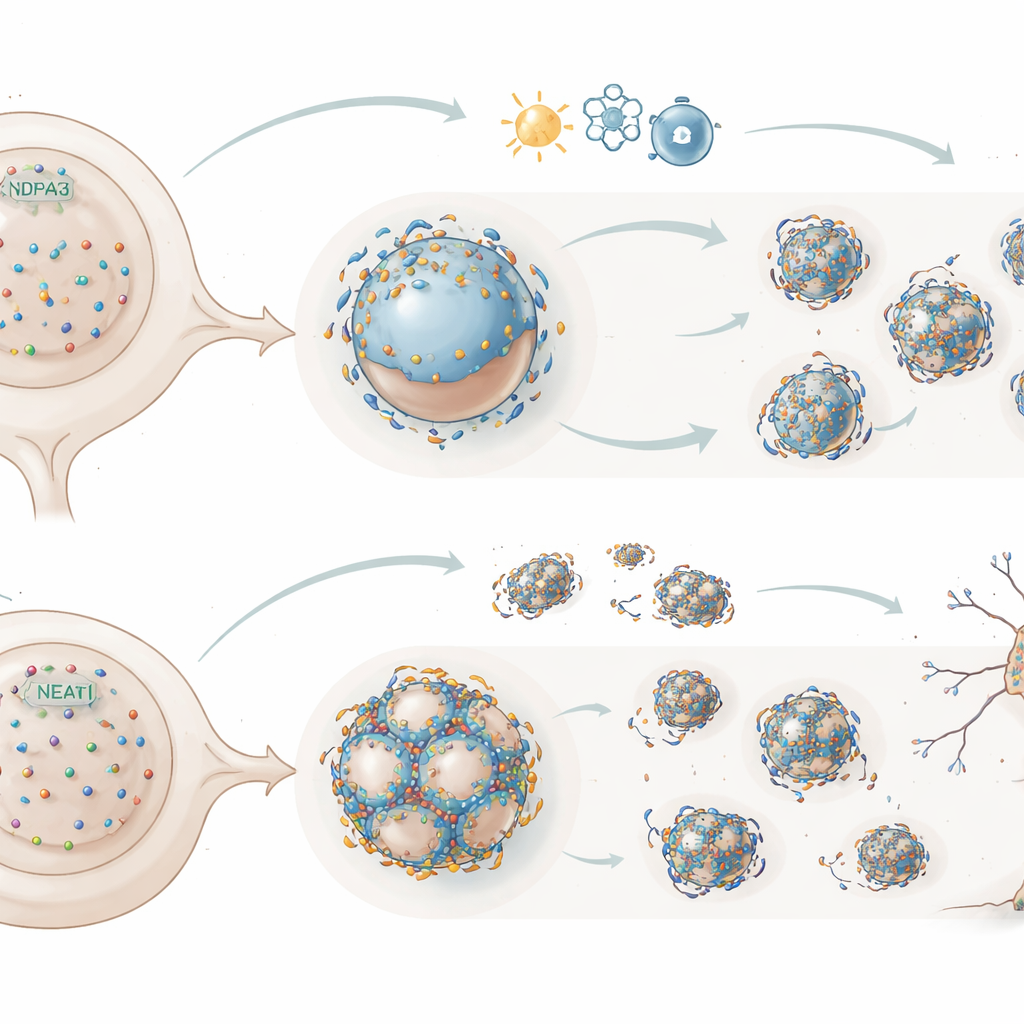
Des gouttelettes dans le noyau qui détectent le stress
Les paraspeckles sont des compartiments sans membrane qui se forment autour de l’ARN NEAT1_2 et d’un ensemble de protéines partenaires. Ils peuvent piéger des molécules régulatrices et ainsi ajuster l’activité de certains gènes, en particulier en situation de stress. Dans la plupart des tissus sains, les paraspeckles sont rares, mais ils apparaissent rapidement lorsque les cellules sont mises à l’épreuve et ont été observés dans les motoneurones de personnes atteintes de SLA. Les auteurs ont cherché à comprendre pourquoi les paraspeckles restent éteints la plupart du temps mais peuvent s’allumer rapidement, et comment TDP-43, une protéine centrale dans la SLA, s’inscrit dans cet interrupteur.
Un frein moléculaire à la formation de gouttelettes
En utilisant des lignées cellulaires humaines et des motoneurones dérivés de cellules souches, les chercheurs ont augmenté ou diminué les niveaux de TDP-43 et ont suivi l’évolution des gouttelettes NEAT1_2 grâce à une imagerie avancée de l’ARN et à la microscopie super‑résolution. Un excès de TDP-43 nucléaire fragmentait les grands paraspeckles en de nombreuses petites particules NEAT1_2, tandis que la réduction de TDP-43 poussait le système dans la direction opposée, favorisant des gouttelettes plus grandes et matures. Des études détaillées sur des mutants ont montré que deux caractéristiques de TDP-43 sont essentielles à cette action de frein : sa capacité à se lier à des segments de NEAT1_2 riches en la séquence à deux lettres « UG », et sa tendance à s’auto‑assembler en polymères. Lorsque l’une ou l’autre propriété était perturbée, TDP-43 ne pouvait plus empêcher la condensation des paraspeckles.
Une lutte d’équilibre entre forces protéiques opposées
L’équipe a également examiné FUS et des protéines « coeur » apparentées des paraspeckles qui favorisent normalement la formation de gouttelettes par ségrégation de phase, un processus comparable à la séparation de l’huile et de l’eau. Dans les cellules et dans des systèmes purifiés, FUS formait facilement des condensats ronds de type liquide, alors que TDP-43 formait des amas plus solides et filamenteux. Lorsque les deux étaient mélangés, même de modestes quantités de TDP-43 fragmentaient et émulsifiaient les gouttes de FUS, les réduisant et empêchant leur fusion. Pourtant, si les chercheurs augmentaient la quantité de FUS, ou d’autres composants centraux comme NONO et SFPQ, ils pouvaient neutraliser cet effet perturbateur et reformer des gouttelettes normales. À l’intérieur des paraspeckles individuels, FUS et ses partenaires se concentraient à l’intérieur, tandis que TDP-43 se disposait en amas irréguliers à faible mobilité à la coque externe, créant des « microphases » distinctes à l’intérieur d’un même condensat. 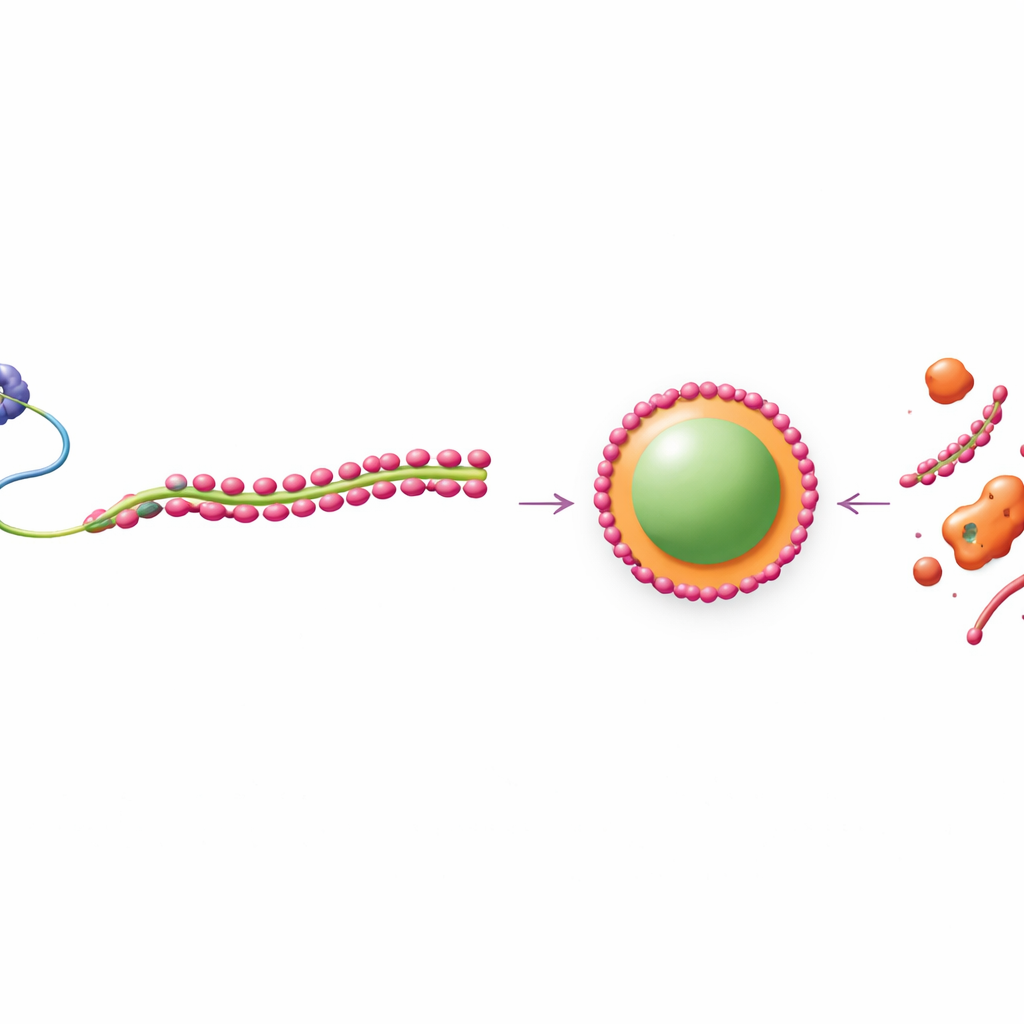
Stress, reprogrammation protectrice et réglage génétique fin
Les cellules soumises à un stress oxydatif ou protéotoxique assemblent de nouveaux condensats nucléaires riches en TDP-43 ailleurs dans le noyau. Les auteurs ont observé qu’à mesure que TDP-43 était attiré dans ces corps induits par le stress, sa liaison à NEAT1_2 se relâchait et les paraspeckles se reformaient, devenant plus dynamiques. Cela suggère que la séquestration de TDP-43 pendant le stress est un mécanisme intégré pour lever le frein et favoriser l’assemblage de gouttelettes protectrices. L’équipe s’est ensuite intéressée aux régions riches en UG de NEAT1_2 elle‑même. Trois blocs répétés UG au milieu de l’ARN recrutent TDP-43 pendant la transcription et sont cruciaux pour le positionner dans les gouttelettes en formation. Un quatrième bloc UG, exceptionnellement long et situé à l’extrémité 3′ de l’ARN, repose à la surface de la goutte et contribue principalement à attirer TDP-43 après l’assemblage, influençant la stabilité des gouttelettes et la rapidité de leur renouvellement.
De l’architecture nucléaire à la survie neuronale
Pour tester l’importance de ce réglage subtil chez les cellules cérébrales, les chercheurs ont supprimé la répétition UG en 3′ de NEAT1_2 chez des neurones humains. Les paraspeckles se formaient toujours, mais ils étaient plus stables lors de l’arrêt de la transcription et plus facilement induits par le stress. De manière frappante, ces motoneurones édités étaient plus résistants à un stress chronique de faible intensité, tandis que les neurones dépourvus complètement de NEAT1_2 et de paraspeckles étaient plus vulnérables. En étendant l’analyse à des milliers de personnes atteintes de SLA, l’équipe a constaté que les individus porteurs de versions particulièrement longues de la répétition UG en 3′ avaient tendance à une survie plus courte. Le travail soutient un modèle selon lequel TDP-43, guidé par l’agencement des répétitions UG sur NEAT1_2, freine normalement la formation des paraspeckles et module leur dynamique. Sous stress, la levée de cette contrainte active un programme cytoprotecteur, mais un excès ou un mauvais positionnement de TDP-43, ou une répétition de queue trop longue, peut surstabiliser la liaison de TDP-43, affaiblir la protection apportée par les paraspeckles et accélérer la neurodégénérescence.
Pourquoi cela compte pour de futures thérapies
Pour les non‑spécialistes, le message clé est que toutes les gouttelettes cellulaires ne sont pas des amas nocifs ; certaines, comme les paraspeckles, semblent faire partie du système de défense intégré de la cellule. TDP-43 agit comme un rhéostat moléculaire qui décide quand et avec quelle intensité ces gouttelettes protectrices se forment, et l’ARN NEAT1_2 encode ce contrôle via le positionnement précis et la longueur de courtes répétitions de séquence. De petites différences génétiques dans ce « bouton de réglage » ARN peuvent influencer la capacité des neurones à supporter un stress prolongé dans la SLA. En apprenant à ajuster ce circuit de condensats — soit en modulant la polymérisation de TDP-43, soit en ciblant les répétitions de NEAT1_2 — il pourrait un jour être possible de renforcer les gouttelettes protectrices de la cellule et de ralentir la progression des maladies neurodégénératives.
Citation: Hodgson, R.E., Huang, WP., Lang, R. et al. Paraspeckle condensation is controlled via TDP-43 polymerization and linked to neuroprotection. Nat Cell Biol 28, 754–770 (2026). https://doi.org/10.1038/s41556-026-01895-y
Mots-clés: paraspeckles, TDP-43, NEAT1, ségrégation de phase, sclérose latérale amyotrophique