Clear Sky Science · en
Paraspeckle condensation is controlled via TDP-43 polymerization and linked to neuroprotection
How tiny nuclear droplets may protect brain cells
Inside each of our cells, especially nerve cells, lie miniature liquidlike droplets called paraspeckles. These structures are built around a long RNA molecule named NEAT1_2 and help cells cope with stress. In many neurodegenerative diseases, another protein, TDP-43, goes awry and clumps, but its everyday job in healthy nuclei has been puzzling. This study uncovers how TDP-43 physically controls the birth and behavior of paraspeckles, and how small changes in this control system may tip the balance between neuron survival and degeneration in disorders such as amyotrophic lateral sclerosis (ALS). 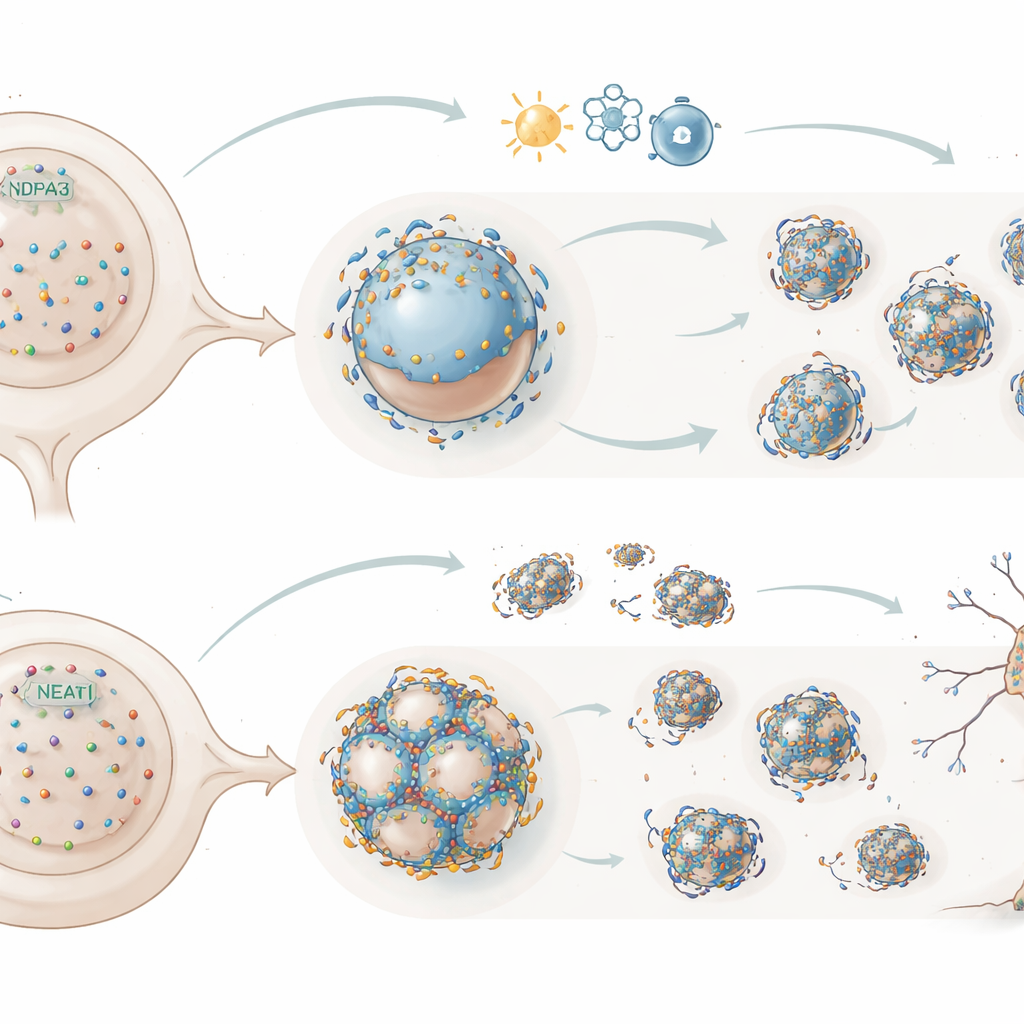
Droplets in the nucleus that sense stress
Paraspeckles are membrane-free compartments that form around the NEAT1_2 RNA and a set of partner proteins. They can trap regulatory molecules and thereby tune which genes are active, particularly during stress. In most healthy tissues, paraspeckles are rare, but they rapidly appear when cells are challenged and have been observed in the motor neurons of people with ALS. The authors set out to understand why paraspeckles stay off most of the time yet can switch on quickly, and how TDP-43, a protein central to ALS, fits into this switch.
A molecular brake on droplet formation
Using human cell lines and stem cell–derived motor neurons, the researchers increased or decreased TDP-43 levels and watched what happened to NEAT1_2 droplets with advanced RNA imaging and super-resolution microscopy. Extra nuclear TDP-43 broke up large paraspeckles into many small NEAT1_2 particles, while reducing TDP-43 pushed the system in the opposite direction, favoring bigger, mature droplets. Detailed mutant studies showed that two features of TDP-43 are essential for this braking action: its ability to bind stretches of NEAT1_2 rich in the two-letter code “UG,” and its tendency to self-assemble into polymers. When either property was disrupted, TDP-43 could no longer prevent paraspeckle condensation.
A tug-of-war between opposing protein forces
The team also examined FUS and related “core” paraspeckle proteins that normally drive droplet formation through phase separation, a process akin to oil separating from water. In cells and in purified systems, FUS readily formed round, liquidlike condensates, whereas TDP-43 formed more solid, filamentous clusters. When both were mixed, even modest amounts of TDP-43 fragmented and emulsified FUS droplets, shrinking them and preventing their fusion. Yet if the researchers boosted FUS, or other core components such as NONO and SFPQ, they could overcome this disruptive effect and re-form normal droplets. Within individual paraspeckles, FUS and its partners concentrated in the interior, while TDP-43 settled into patchy, low-mobility clusters at the outer shell, creating distinct “microphases” inside one condensate. 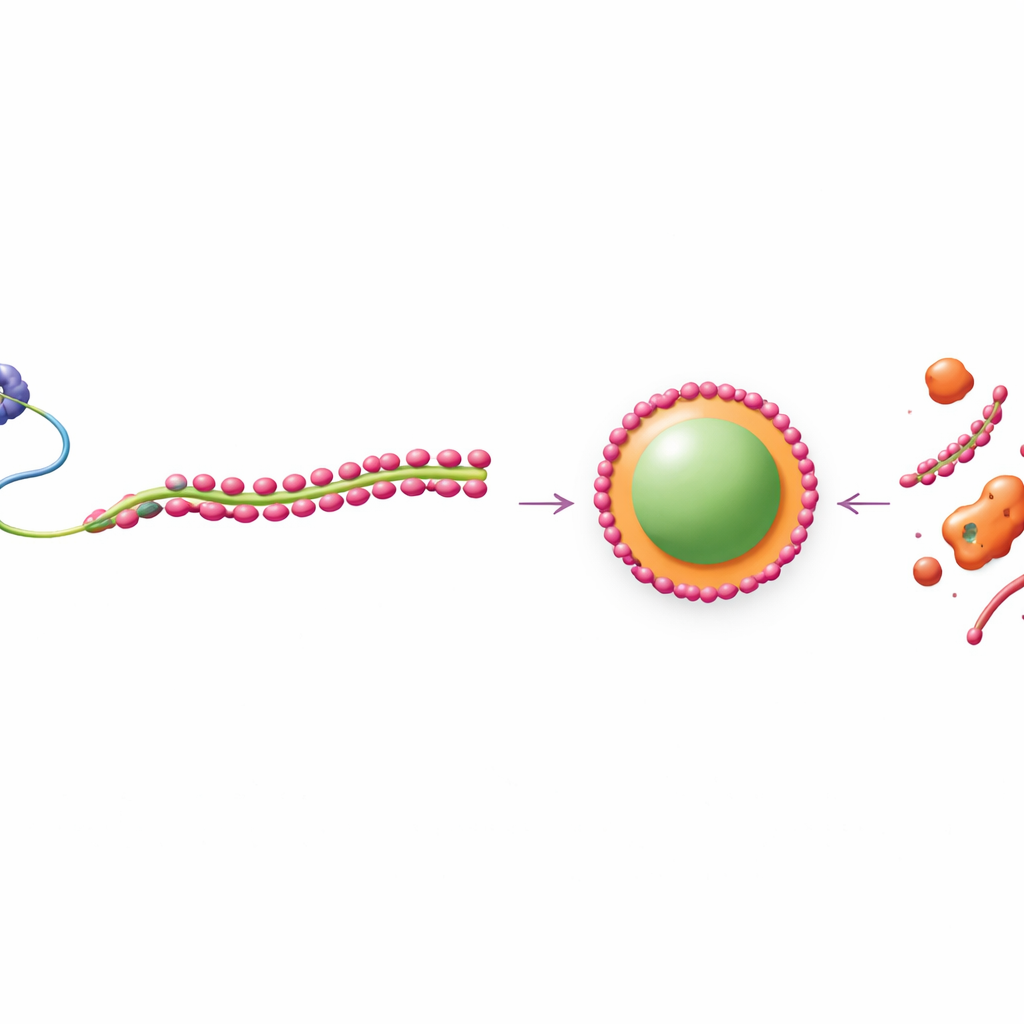
Stress, protective rewiring, and genetic fine-tuning
Cells under oxidative or proteotoxic stress assemble new TDP-43–rich nuclear condensates elsewhere in the nucleus. The authors observed that as TDP-43 was drawn into these stress-induced bodies, its grip on NEAT1_2 relaxed and paraspeckles re-formed, becoming more dynamic. This suggests that sequestration of TDP-43 during stress is a built-in way to lift the brake and promote protective droplet assembly. The team then zoomed in on the UG-rich regions of NEAT1_2 itself. Three UG repeat blocks in the middle of the RNA recruit TDP-43 during transcription and are crucial for placing it into the forming droplets. A fourth, unusually long UG block at the RNA’s tail-end sits on the droplet surface and mainly helps draw in TDP-43 after assembly, influencing how stable the droplets remain and how quickly they turn over.
From nuclear architecture to neuron survival
To test how this subtle tuning matters for brain cells, the researchers removed the 3′-end UG repeat from NEAT1_2 in human neurons. Paraspeckles still formed, but they were more stable during transcriptional shutdown and more readily induced by stress. Strikingly, these edited motor neurons were more resistant to chronic low-level stress, while neurons completely lacking NEAT1_2 and paraspeckles were more vulnerable. Extending the work to thousands of people with ALS, the team found that individuals carrying especially long versions of the 3′ UG repeat tended to have shorter survival. The work supports a model in which TDP-43, guided by the layout of UG repeats on NEAT1_2, normally restrains paraspeckle formation and modulates their movement. Under stress, releasing this restraint boosts a cytoprotective program, but excessive or misplaced TDP-43, or an overly long tail repeat, may over-stabilize TDP-43 binding, weaken paraspeckle-based protection, and accelerate neurodegeneration.
Why this matters for future therapies
For non-specialists, the key message is that not all cellular droplets are harmful clumps; some, like paraspeckles, appear to be part of the cell’s built-in defense system. TDP-43 acts as a molecular rheostat that decides when and how strongly these protective droplets assemble, and the NEAT1_2 RNA encodes this control through the precise positioning and length of short sequence repeats. Small genetic differences in this RNA “tuning knob” can influence how well neurons weather long-term stress in ALS. By learning how to adjust this condensate circuitry—either by modulating TDP-43 polymerization or by targeting NEAT1_2 repeats—it may one day be possible to enhance the cell’s own protective droplets and slow the course of neurodegenerative disease.
Citation: Hodgson, R.E., Huang, WP., Lang, R. et al. Paraspeckle condensation is controlled via TDP-43 polymerization and linked to neuroprotection. Nat Cell Biol 28, 754–770 (2026). https://doi.org/10.1038/s41556-026-01895-y
Keywords: paraspeckles, TDP-43, NEAT1, phase separation, amyotrophic lateral sclerosis