Clear Sky Science · fr
VEGFR2 est requis pour l’angiogenèse lymphatique de type sprouting médiée par VEGF-C–VEGFR3–PI3Kα
Pourquoi de minuscules vaisseaux de drainage comptent
Les vaisseaux lymphatiques forment un système de drainage caché qui élimine l’excès de liquide, les lipides et les cellules immunitaires de nos tissus. Lorsque ce réseau se développe insuffisamment, un gonflement et une mauvaise cicatrisation peuvent en résulter ; lorsqu’il se développe excessivement, il peut alimenter l’inflammation chronique et favoriser la dissémination des cancers. Cette étude explore comment deux « antennes » moléculaires proches sur les cellules lymphatiques coopèrent pour guider le bourgeonnement contrôlé de nouveaux vaisseaux lymphatiques, un processus appelé lymphangiogenèse, avec des implications potentielles pour le traitement du lymphœdème, de l’inflammation chronique et du cancer.
Deux interrupteurs moléculaires sur les vaisseaux lymphatiques
Les vaisseaux lymphatiques sont tapissés de cellules spécialisées qui détectent les signaux de croissance dans leur environnement. Un signal clé, nommé VEGF-C, est connu pour stimuler la formation de nouveaux vaisseaux lymphatiques en se liant à un récepteur appelé VEGFR3 sur ces cellules. Un récepteur apparenté, VEGFR2, était considéré comme important surtout pour les vaisseaux sanguins plutôt que lymphatiques. Des études animales antérieures utilisant des outils génétiques moins précis suggéraient que les vaisseaux lymphatiques pouvaient croître sans beaucoup de contribution de VEGFR2, alors que des expériences en culture cellulaire laissaient entendre que VEGFR2 était puissant pour promouvoir le mouvement et la survie cellulaires. Ce décalage posait une question centrale : VEGFR2 compte‑t‑il vraiment pour le bourgeonnement des vaisseaux lymphatiques in vivo, et si oui, comment ?
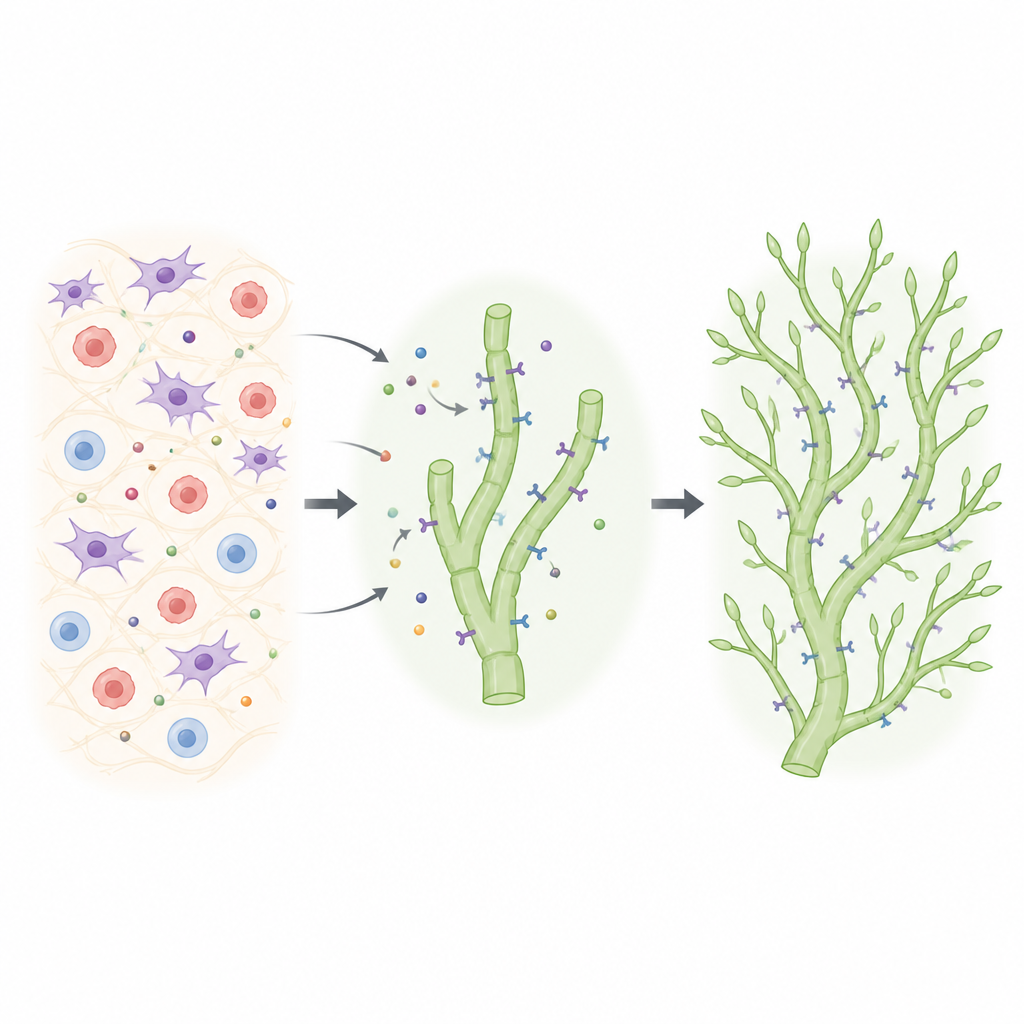
Révéler une dépendance cachée à VEGFR2
Les chercheurs ont utilisé une génétique murine affinée pour inactiver VEGFR2 spécifiquement dans les cellules lymphatiques à des moments choisis, tout en suivant quelles cellules avaient réellement perdu le récepteur. Ils ont observé la formation des réseaux lymphatiques dans la peau de l’oreille, où les vaisseaux bourgeonnent normalement à partir de troncs profonds pour construire un maillage capillaire délicat après la naissance. Lorsque VEGFR2 était supprimé seulement dans certaines cellules lymphatiques, ces cellules étaient systématiquement désavantagées : les fronts de bourgeonnement devenaient dominés par des cellules ayant conservé VEGFR2. Ce n’est que lorsque l’équipe obtint une suppression très efficace et continue qu’un défaut net apparut : le réseau capillaire devint clairsemé, avec moins de branches et d’extrémités. Fait frappant, lorsque VEGFR2 et VEGFR3 furent supprimés simultanément chez des nouveau‑nés, le réseau lymphatique dermique ne se forma pas, révélant que les deux récepteurs sont nécessaires à la survie et à la croissance appropriée de ces vaisseaux.
Séparer l’élargissement des vaisseaux du bourgeonnement
Pour analyser comment chaque récepteur influence le comportement vasculaire dans les tissus adultes, l’équipe augmenta les niveaux de VEGF-C dans la peau via des vecteurs viraux tout en supprimant sélectivement VEGFR2, VEGFR3 ou une enzyme clé en aval appelée PI3Kα dans les cellules lymphatiques. Lorsque VEGFR3 manquait seul, VEGF-C ne pouvait plus du tout déclencher la croissance lymphatique. Lorsque seul VEGFR2 était absent, les vaisseaux lymphatiques s’agrandissaient encore et leurs cellules se divisaient, mais l’explosion habituelle de nouveaux bourgeons et ramifications latérales disparaissait. Ce schéma reproduisait étroitement ce qui se produisait lorsque VEGF-C était conçu pour n’activer que VEGFR3 et pas VEGFR2. En revanche, la suppression de PI3Kα bloquait complètement à la fois l’élargissement et le bourgeonnement. Ensemble, ces résultats suggèrent que VEGFR3 suffit à stimuler la prolifération cellulaire, tandis qu’un partenariat entre VEGFR2, VEGFR3 et PI3Kα est nécessaire pour convertir cette expansion cellulaire en nouvelles branches vasculaires.
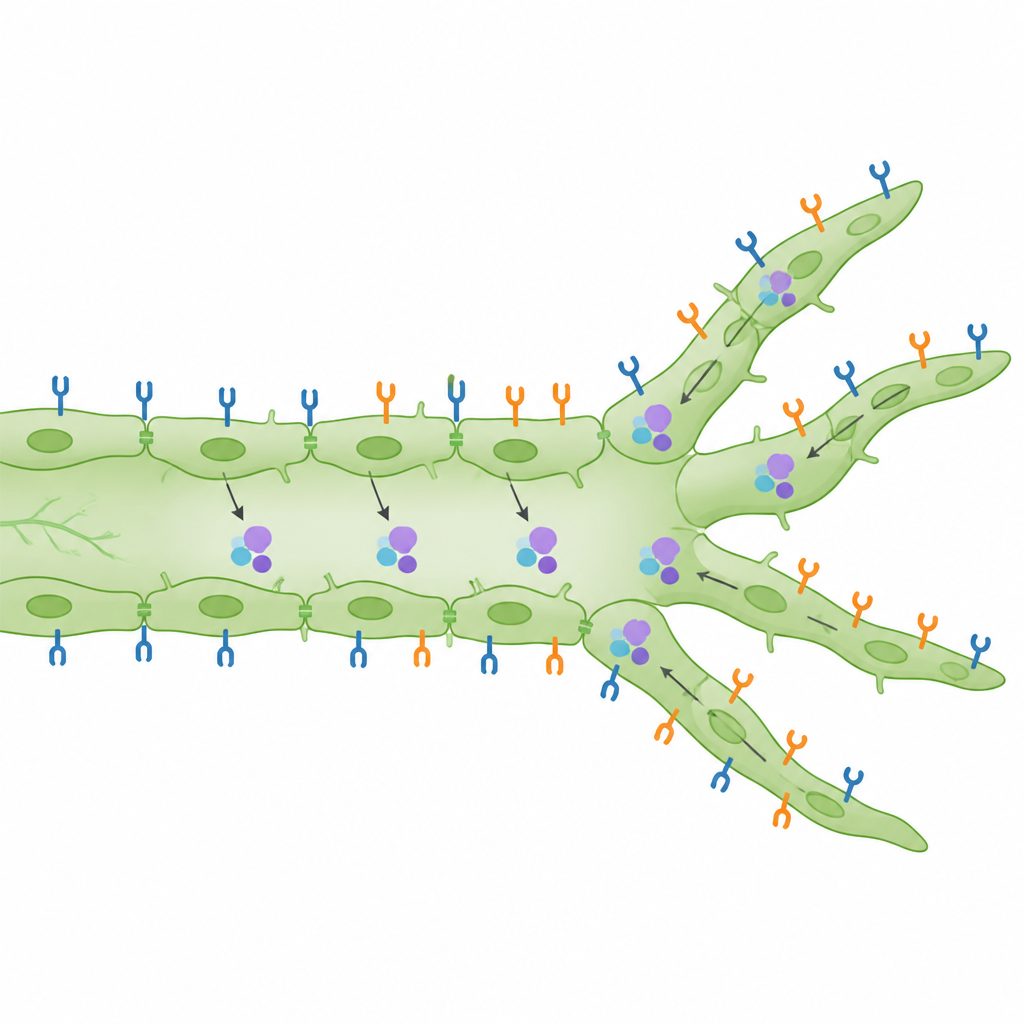
Comment les récepteurs sont organisés et activés
Au‑delà du dénombrement des branches, les scientifiques ont examiné où les récepteurs se situent à la surface cellulaire et s’ils sont activés. Ils ont constaté que, tant chez l’embryon en développement que dans la peau adulte, VEGFR2 est étonnamment abondant à la surface des cellules lymphatiques comparé aux cellules des vaisseaux sanguins voisins. Lorsque les niveaux de VEGF-C augmentaient, les protéines VEGFR3 se déplaçaient vers l’intérieur des cellules, cohérent avec une signalisation active, tandis que les niveaux de VEGFR2 à la surface des cellules lymphatiques augmentaient d’une façon dépendante de PI3Kα. Un test de proximité sensible dans la peau embryonnaire révéla que VEGFR2 et VEGFR3 sont non seulement actifs, mais souvent très proches l’un de l’autre, ce qui appuie l’idée qu’ils peuvent former des paires mixtes envoyant des signaux distincts. Dans des expériences de cicatrisation par plaie utilisant un modèle de perforation de l’oreille, la perte de VEGFR2 réduisit spécifiquement le nombre de pointes acérées et en bourgeon qui repoussaient dans la zone lésée, tandis que la perte de VEGFR3 eut un effet plus marqué sur la quantité totale de nouveau tissu lymphatique.
Équilibrer croissance et ramification dans le réseau lymphatique
Dans l’ensemble, l’étude propose que VEGF-C stimule d’abord VEGFR3 pour faire diviser les cellules lymphatiques, puis, via PI3Kα et une augmentation de VEGFR2 à la surface cellulaire, oriente la signalisation vers des paires VEGFR2–VEGFR3 qui favorisent la formation de bourgeons. Cette passation coordonnée aide à garantir qu’il y a suffisamment de cellules pour former de nouvelles branches, sans excès ni insuffisance qui rendraient le réseau dysfonctionnel. Pour le lecteur non spécialiste, l’idée principale est que deux interrupteurs moléculaires similaires, auparavant considérés comme jouant des rôles très différents, coopèrent en réalité étroitement pour façonner le bourgeonnement des nouveaux vaisseaux lymphatiques. Comprendre ce partenariat pourrait orienter de futures tentatives pour stimuler la croissance lymphatique, par exemple pour améliorer la cicatrisation, ou la freiner lorsque son expansion excessive contribue à la maladie.
Citation: Schoofs, H., Zhang, Y., Ortsäter, H. et al. VEGFR2 is required for VEGF-C–VEGFR3–PI3Kα-mediated sprouting lymphangiogenesis. Nat Commun 17, 4380 (2026). https://doi.org/10.1038/s41467-026-73013-3
Mots-clés: vaisseaux lymphatiques, VEGF-C, VEGFR2, bourgeonnement, PI3K alpha