Clear Sky Science · en
VEGFR2 is required for VEGF-C–VEGFR3–PI3Kα-mediated sprouting lymphangiogenesis
Why tiny drainage vessels matter
Lymphatic vessels form a hidden drainage system that clears excess fluid, fats and immune cells from our tissues. When this network grows too little, swelling and poor healing can follow; when it grows too much, it can fuel chronic inflammation and help cancers spread. This study digs into how two closely related molecular "antennae" on lymphatic cells work together to guide the controlled sprouting of new lymphatic vessels, a process called lymphangiogenesis, with potential implications for treating lymphedema, chronic inflammation and cancer.
Two molecular switches on lymphatic vessels
Lymphatic vessels are lined by specialized cells that sense growth signals in their environment. One key signal, called VEGF-C, is known to drive the growth of new lymphatic vessels by binding to a receptor named VEGFR3 on these cells. A sister receptor, VEGFR2, was thought to be important mainly in blood vessels rather than lymphatic ones. Earlier animal studies using less precise genetic tools suggested that lymphatic vessels could grow without much help from VEGFR2, even though cell culture experiments hinted that VEGFR2 was powerful at driving cell movement and survival. This mismatch raised a central question: does VEGFR2 truly matter for lymphatic vessel sprouting inside a living body, and if so, how?
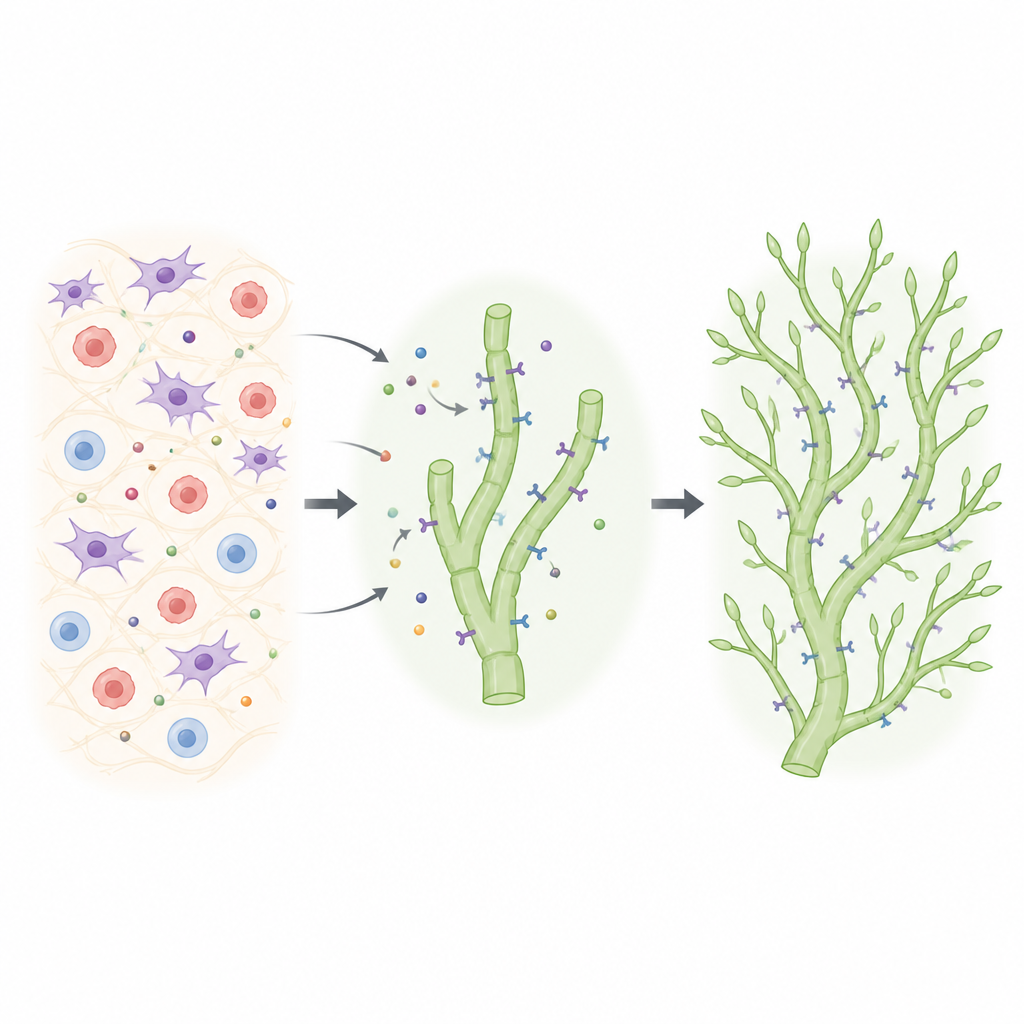
Revealing a hidden dependency on VEGFR2
The researchers used refined mouse genetics to turn VEGFR2 off specifically in lymphatic cells at chosen times, while also tracking which cells had truly lost the receptor. They followed how lymphatic networks form in the skin of the ear, where vessels normally sprout from deeper trunks to build a delicate capillary web after birth. When VEGFR2 was deleted only in some lymphatic cells, those cells were repeatedly outcompeted: the sprouting fronts became dominated by cells that had managed to keep VEGFR2. Only when the team achieved highly efficient and continuous deletion did a clear defect emerge: the capillary network became sparse, with fewer branches and tips. Strikingly, when both VEGFR2 and VEGFR3 were removed together in newborn mice, the dermal lymphatic network failed to form, revealing that both receptors are required for survival and proper growth of these vessels.
Separating vessel enlargement from sprouting
To probe how each receptor shapes vessel behavior in adult tissues, the team boosted VEGF-C levels in the skin using viral vectors while selectively removing VEGFR2, VEGFR3 or a key downstream enzyme called PI3Kα in lymphatic cells. When VEGFR3 alone was missing, VEGF-C could no longer trigger lymphatic growth at all. When only VEGFR2 was removed, lymphatic vessels still enlarged and their cells still divided, but the usual explosion of new sprouts and side branches was lost. This pattern closely mimicked what happened when VEGF-C was engineered to activate only VEGFR3 but not VEGFR2. In contrast, deleting PI3Kα completely blocked both enlargement and sprouting. Together, these findings suggest that VEGFR3 is sufficient to drive cell proliferation, while a partnership between VEGFR2, VEGFR3 and PI3Kα is necessary to convert that cell expansion into new vessel branches.
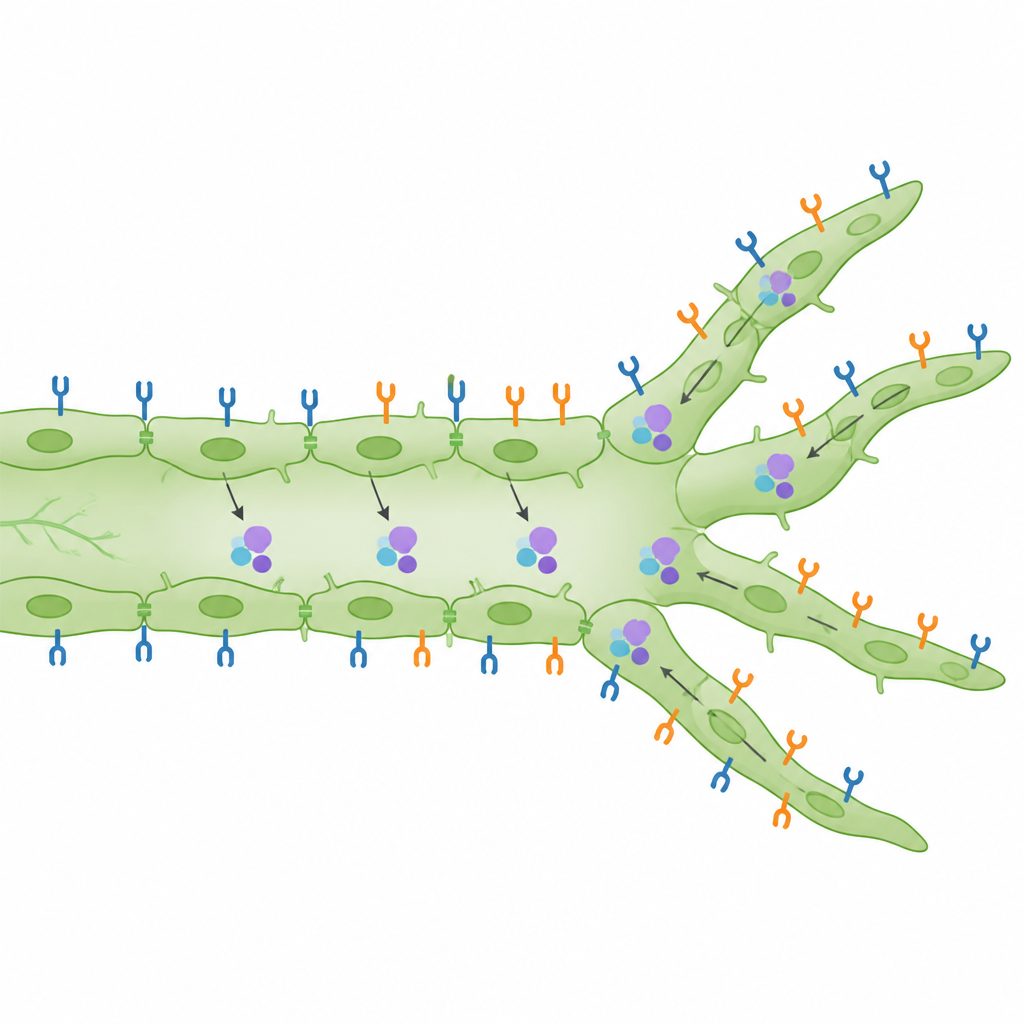
How the receptors are arranged and activated
Beyond counting branches, the scientists examined where the receptors sit on the cell surface and whether they are switched on. They found that, in both developing embryos and adult skin, VEGFR2 is surprisingly abundant on the surface of lymphatic cells compared with neighboring blood vessel cells. When VEGF-C levels were raised, VEGFR3 proteins moved inward inside the cells, consistent with active signaling, while VEGFR2 levels at the surface of lymphatic cells increased in a manner that depended on PI3Kα. A sensitive proximity assay in embryonic skin revealed that VEGFR2 and VEGFR3 are not only active, but often lie very close together, supporting the idea that they can form mixed pairs that send distinct signals. In wound-healing experiments using an ear punch model, loss of VEGFR2 specifically reduced the number of sharp, sprouting tips that regrew into the injured zone, while VEGFR3 loss had a stronger effect on the overall amount of new lymphatic tissue.
Balancing growth and branching in the lymphatic network
Overall, the study proposes that VEGF-C first stimulates VEGFR3 to make lymphatic cells divide, then, through PI3Kα and an increase in VEGFR2 at the cell surface, shifts signaling toward VEGFR2–VEGFR3 pairs that favor sprout formation. This coordinated handoff helps ensure that there are enough cells to build new branches, but not so much or so little growth that the network becomes dysfunctional. For a lay reader, the key takeaway is that two similar molecular switches, once thought to play very different roles, actually cooperate closely to shape how new lymphatic vessels sprout. Understanding this partnership could guide future attempts to either boost lymphatic growth, for example to improve wound healing, or rein it in when excessive lymphatic expansion contributes to disease.
Citation: Schoofs, H., Zhang, Y., Ortsäter, H. et al. VEGFR2 is required for VEGF-C–VEGFR3–PI3Kα-mediated sprouting lymphangiogenesis. Nat Commun 17, 4380 (2026). https://doi.org/10.1038/s41467-026-73013-3
Keywords: lymphatic vessels, VEGF-C, VEGFR2, sprouting, PI3K alpha