Clear Sky Science · fr
Un modèle multifacette de KERP2 d’Entamoeba histolytica régulant l’expression génique et les réponses des cellules hôtes
Comment un minuscule parasite peut miner nos défenses intestinales
L’amibiase est une infection intestinale qui touche des millions de personnes dans le monde, provoquant parfois des diarrhées sévères et des lésions intestinales potentiellement mortelles. Cette étude examine comment une seule protéine du parasite, appelée KERP2, peut à la fois réguler les gènes du parasite et perturber les cellules qui tapissent nos intestins, offrant une fenêtre sur la façon dont les envahisseurs microscopiques déjouent nos défenses.
Un examen approfondi d’une amibe envahissant l’intestin
Le parasite Entamoeba histolytica vit à la surface de la paroi intestinale, où il peut provoquer une inflammation et endommager les tissus. Contrairement aux microbes qui s’abritent à l’intérieur de nos cellules, cette amibe reste à l’extérieur mais parvient néanmoins à manipuler la biologie de l’hôte. Des travaux antérieurs avaient identifié une famille de protéines parasitaires appelées KERP proches de la bordure en brosse, les microvillosités des cellules intestinales. L’une d’elles, KERP2, paraissait particulièrement intéressante car elle est conservée chez des amibes apparentées et associée à la gravité de la maladie. Curieusement, KERP2 semblait être ciblée vers le noyau du parasite, même si on la retrouvait également dans des fractions associées à la membrane à la surface cellulaire.
Une protéine aux formes changeantes à l’intérieur du parasite
Grâce à des comparaisons de séquences et des prédictions structurelles informatiques, les auteurs montrent que KERP2 possède des caractéristiques rappelant une protéine de la chromatine appelée DEK, connue dans d’autres organismes pour remodeler la compaction et la lecture de l’ADN. KERP2 contient un module de type SAP qui préfère l’ADN riche en A et T, et une queue en coiled-coil qui porte un signal d’adressage nucléaire. Des expériences avec des versions marquées de KERP2 révèlent que la protéine pleine longueur s’accumule dans le noyau du parasite, notamment dans les régions denses en ADN, tandis qu’une version privée de la queue coiled-coil reste majoritairement dans le cytoplasme. Dans des essais in vitro, KERP2 se lie fortement à l’ADN riche en AT mais pas à l’ADN riche en GC, et semble fléchir ou compacter l’ADN plutôt que reconnaître une séquence précise. Ensemble, ces éléments décrivent KERP2 comme un auxiliaire associé à la chromatine qui affine l’expression de groupes de gènes, plutôt qu’un interrupteur on–off classique.
Ajuster les armes du parasite
Pour déterminer le rôle de KERP2 pour le parasite, l’équipe a réduit sa production par silencement génique. Les parasites dépourvus de KERP2 se multipliaient normalement, mais leur profil d’expression génique changeait. De nombreux gènes liés à la pathogénicité amibienne, notamment ceux codant des protéases à cystéine et des peptides formant des pores, étaient plus actifs, tout comme des gènes impliqués dans le métabolisme du soufre et des acides aminés. Des tests enzymatiques directs ont confirmé que l’activité des protéases à cystéine, principal moteur des lésions tissulaires, augmentait lors du knockdown de KERP2 et diminuait lorsqu’on surexprimait KERP2. Des études d’interaction ont également montré que KERP2 s’associe à des facteurs de transport nucléaire, des protéines liant l’ARN et l’ADN, des composants ribosomiques et des protéines de trafic comme Rab11B, suggérant que KERP2 se situe au carrefour de la régulation génique et des voies sécrétoires.
Du parasite aux cellules humaines
L’histoire ne s’arrête pas à l’intérieur du parasite. Lorsqu’elles entrent en contact avec des lignées cellulaires intestinales humaines ou un modèle tridimensionnel de cryptes humaines, on peut détecter KERP2 à l’intérieur des cellules hôtes. L’imagerie et la fractionation montrent des signaux ponctués de KERP2 dans le cytoplasme de l’hôte et près des microvillosités, et de faibles quantités même dans le noyau, bien qu’un rôle direct dans le contrôle de l’ADN de l’hôte ne soit pas encore démontré. La protéine KERP2 purifiée peut, à elle seule, pénétrer dans les cellules intestinales par un processus dépendant de l’énergie ressemblant à l’endocytose et y rester au moins deux jours. Une fois présente dans les cellules hôtes, KERP2 s’associe à des protéines qui gouvernent le cytosquelette d’actine, les jonctions cellulaires et la signalisation. Le profilage génique de l’hôte révèle des modifications des réponses au stress, du métabolisme et des voies liées à la division cellulaire et à l’organisation structurale.
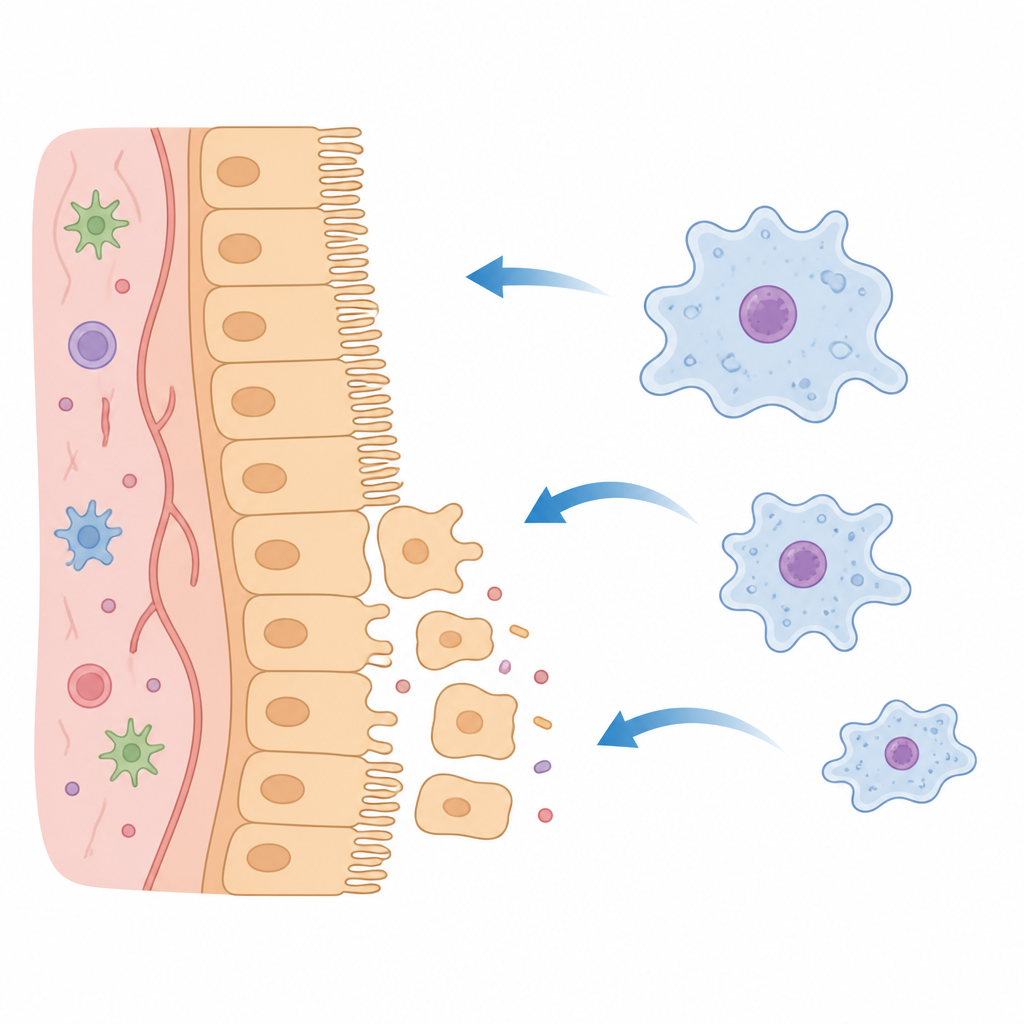
Reprogrammer la forme cellulaire et la force de la barrière
Au plan fonctionnel, KERP2 modifie le comportement des cellules intestinales et leur cohésion. Les cellules exposées à KERP2, que ce soit via des parasites vivants ou l’ajout de protéine purifiée, montrent une augmentation de la synthèse d’ADN, suggérant une incitation vers l’activité du cycle cellulaire. Leurs fibres d’actine se réorganisent, les cellules deviennent plus allongées et l’anneau dense d’actine habituel aux bords cellulaires s’affaiblit. Dans des tests de cicatrisation visant à suivre la vitesse de fermeture d’un feuillet cellulaire, KERP2 ralentit le mouvement collectif lorsque la division cellulaire nouvelle est limitée. Des mesures de la résistance électrique à travers des couches cellulaires et le suivi de molécules fluorescentes traversant la barrière révèlent que KERP2 peut réduire l’étanchéité ionique et, dans certains cas, augmenter la fuite de molécules de plus grande taille via les jonctions. Fait intéressant, lorsque le parasite est privé de KERP2, il compense par une activité protéasique plus élevée, provoquant des chutes importantes de la résistance électrique mais pas toujours des fuites de grosses molécules, ce qui implique que le cytosquelette de l’hôte peut parfois se resserrer pour contrer les dommages.
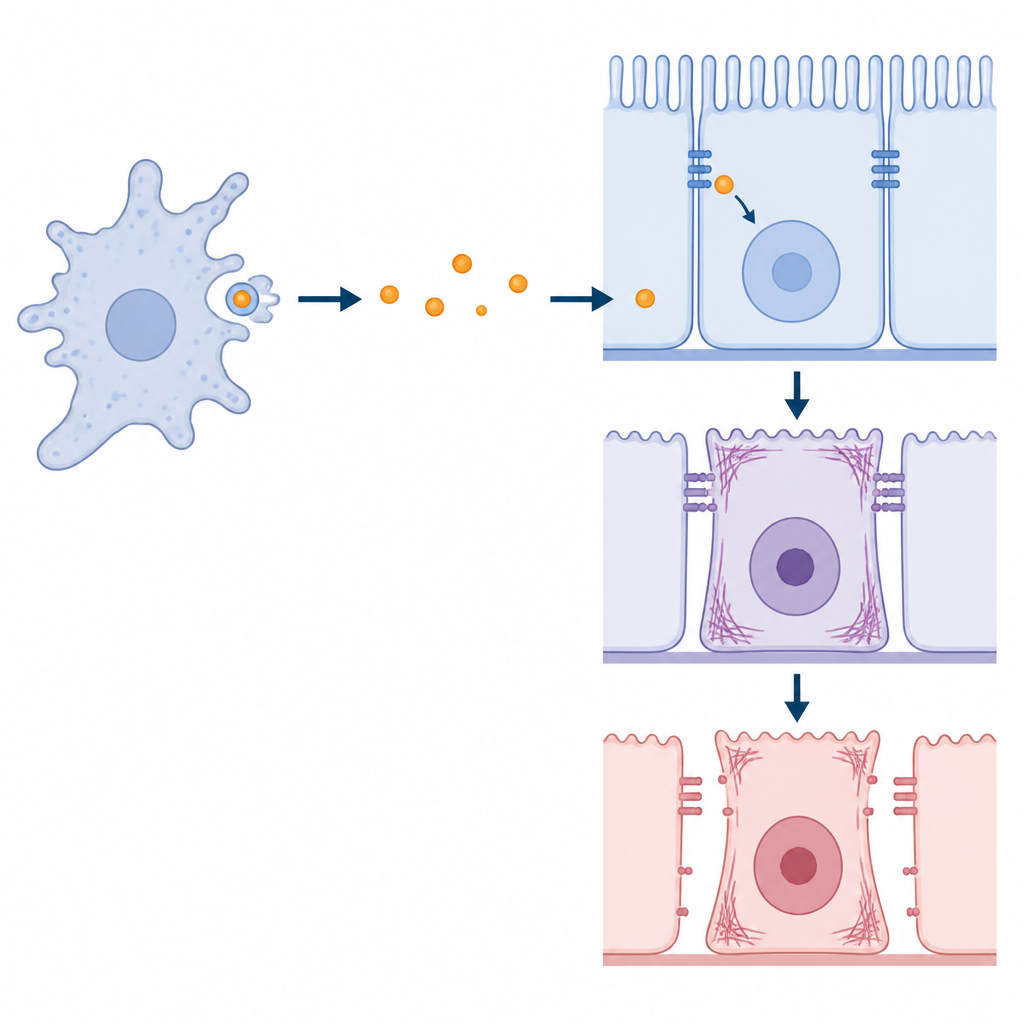
Ce que cela signifie pour la compréhension de l’infection
Ces travaux suggèrent que KERP2 est un outil à double usage : à l’intérieur d’Entamoeba, il aide à maintenir l’équilibre des gènes de virulence et du métabolisme, tandis que lors du contact avec la muqueuse intestinale il peut être transféré dans les cellules hôtes pour ajuster leur structure, leur croissance et leurs propriétés de barrière. Plutôt que d’agir uniquement comme une toxine, KERP2 semble affiner le degré de rigidité ou de perméabilité de la surface intestinale, aidant potentiellement le parasite à s’adapter à différents environnements de l’hôte. Bien que des recherches supplémentaires soient nécessaires, notamment dans des modèles animaux, l’étude offre une vision plus large de la manière dont des parasites extracellulaires peuvent utiliser des protéines multitâches pour coordonner leurs programmes géniques avec un remodelage subtil des tissus hôtes.
Citation: Peng, R., Santos, H.J. & Nozaki, T. A multifaceted model of Entamoeba histolytica KERP2 regulating gene expression and host cell responses. Nat Commun 17, 4433 (2026). https://doi.org/10.1038/s41467-026-70847-9
Mots-clés: Entamoeba histolytica, amibiase, épithélium intestinal, interaction hôte-pathogène, virulence parasitaire