Clear Sky Science · fr
États TRAAK échangés et non échangés coexistent dans les membranes à un ratio influencé par la température
Pourquoi cette petite porte dans les cellules nerveuses compte
Chaque pensée, contact et battement de cœur dépend des signaux électriques qui parcourent nos cellules. Un acteur clé dans la modulation de ces signaux est une famille de protéines « canaux de fuite » qui laissent les ions potassium traverser la membrane cellulaire. Un membre, nommé TRAAK, participe à définir l’excitabilité des cellules nerveuses et intervient même dans la détection de la température et des forces mécaniques. Cette étude révèle comment un petit « capuchon » mobile situé au‑dessus de TRAAK se comporte dans des membranes cellulaires réelles, comment la chaleur influence sa configuration préférée, et comment les lipides environnants forment un voisinage spécifique qui module la fonction du canal. 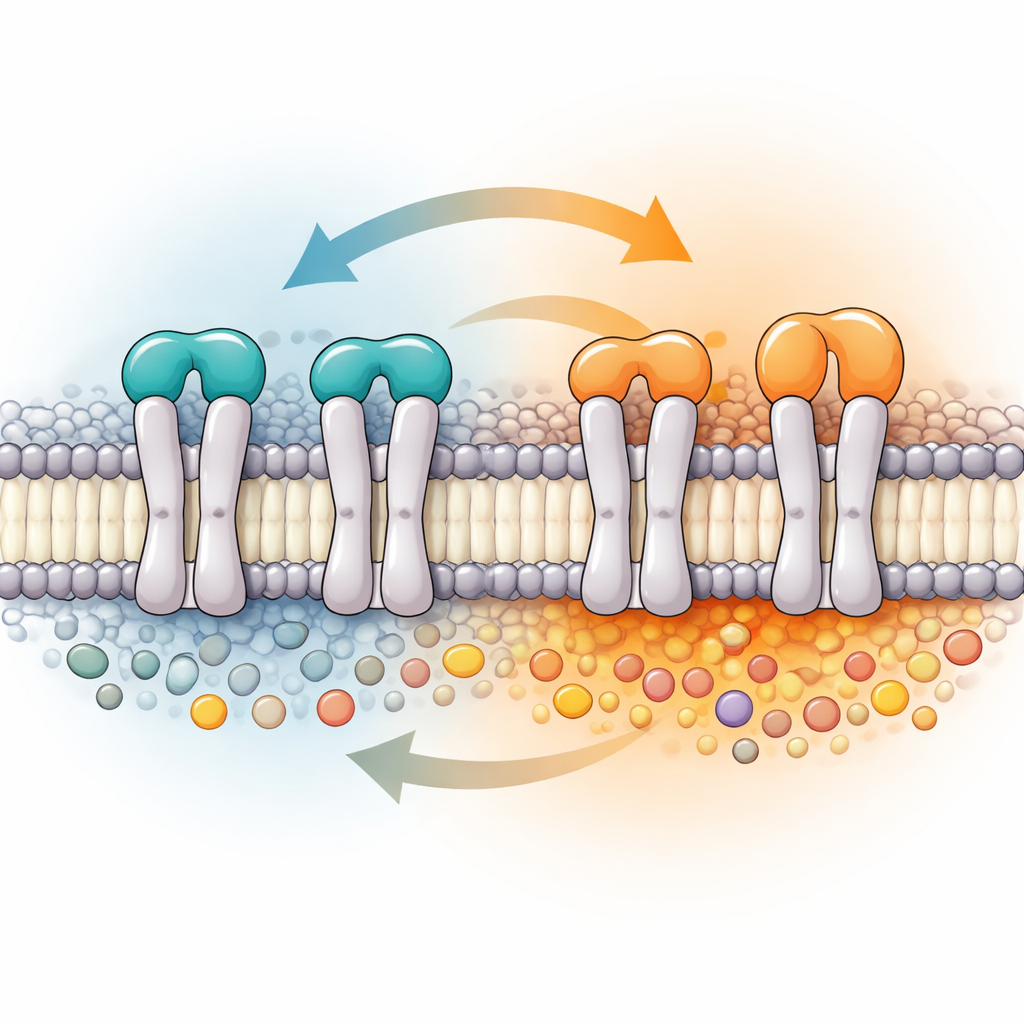
Deux façons d’agencer une porte microscopique
TRAAK appartient à un groupe dit de canaux potassiques à deux domaines de pore qui forment des voies de fuite de fond, maintenant l’équilibre électrique de nombreux types cellulaires, y compris dans le cerveau et le cœur. Contrairement aux canaux potassiques plus familiers à quatre sous‑unités, TRAAK est constitué de deux grandes sous‑unités et porte un capuchon distinctif qui dépasse de la membrane et divise le trajet des ions. Des structures à haute résolution antérieures ont montré que ce capuchon peut adopter deux agencements : une configuration « échangée » où des éléments d’une sous‑unité croisent pour contacter l’autre, et une configuration « non‑échangée » où les parties de chaque sous‑unité restent avec leur partenaire d’origine. Jusqu’à présent, personne ne savait si les deux formes coexistaient réellement dans les membranes naturelles, quelle était la fréquence de chacune, ni ce qui fait basculer l’équilibre entre elles.
Observer des pièces mobiles avec des règles magnétiques
Pour répondre à ces questions, les auteurs ont utilisé une technique magnétique spécialisée appelée résonance paramagnétique électronique dipolaire à impulsions, qui peut mesurer des distances de l’ordre du milliardième de mètre entre de petites étiquettes magnétiques. Ils ont conçu la TRAAK humaine de sorte qu’une seule des deux sous‑unités porte une paire de ces étiquettes à des positions choisies avec soin reliant le capuchon et la région transmembranaire. Ainsi, ils ont pu distinguer les deux formes du capuchon parce que chacune produit une distance étiquette‑étiquette différente, un peu comme mesurer l’écartement entre deux charnières dans des conceptions alternatives de portes. Ils ont aussi intégré TRAAK dans des fragments membranaires maintenus sans détergents agressifs, préservant les lipides naturels voisins et garantissant que la protéine en pleine longueur—avec toutes ses parties régulatrices—restait fonctionnelle. 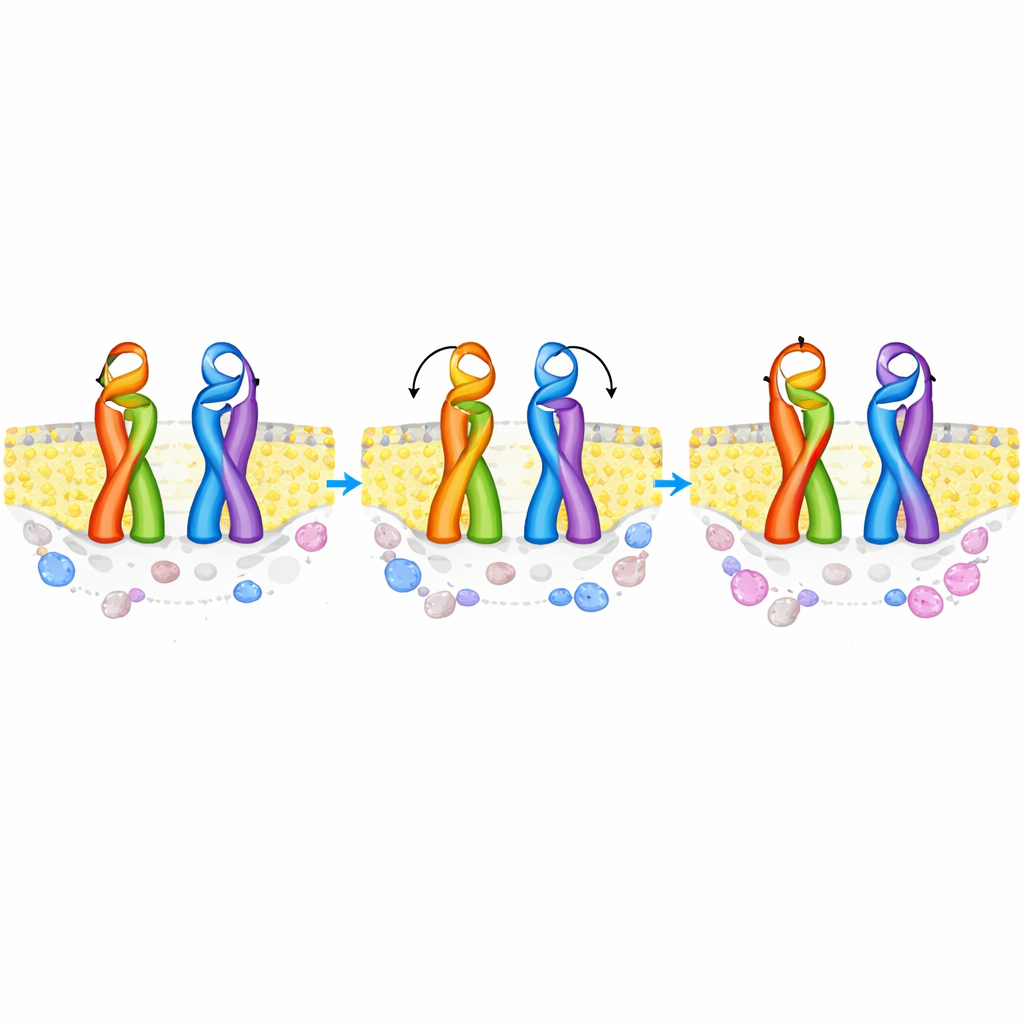
Deux formes partagent la scène, et la chaleur change le scénario
Les mesures de distance ont révélé deux populations distinctes correspondant aux prédictions informatiques pour les capuchons échangés et non‑échangés, prouvant que les deux formes coexistent dans le même ensemble membranaire. À une température proche de la pièce, 19 °C, la forme échangée constituait une majorité modeste des canaux, la forme non‑échangée représentant néanmoins une minorité substantielle. Lorsque les chercheurs ont préparé les échantillons membranaires à 40 °C, l’équilibre a davantage basculé en faveur de l’état échangé, et la configuration non‑échangée est devenue plus rare. Des simulations informatiques de TRAAK dans des membranes modèles suggèrent que l’arrangement échangé est intrinsèquement plus stable et moins sensible à la température, tandis que la forme non‑échangée devient énergétiquement moins favorable à mesure que la membrane et ses lipides se réorganisent sous l’effet de la chaleur.
Un voisinage lipidique sur mesure autour de TRAAK
Au‑delà des changements de forme, TRAAK s’est aussi montré sélectif quant à la compagnie qu’il choisit dans la membrane. En analysant les lipides restés associés à TRAAK lors d’une purification sans détergent, l’équipe a trouvé un enrichissement marqué en phosphatidylinositols et en leurs variantes signalétiques—des lipides connus pour influencer de nombreux canaux ioniques—ainsi qu’en autres espèces chargées négativement. De manière surprenante, le lipide membranaire le plus courant, la phosphatidylcholine, était quasiment absent des environs immédiats de TRAAK, alors qu’il domine la membrane de la cellule hôte. Lors de la reconstitution dans des vésicules artificielles, certains des lipides enrichis ont activé le courant TRAAK, tandis que d’autres l’ont atténué ou n’ont eu que peu d’effet, montrant que le microdomaine lipidique privilégié par le canal n’est pas seulement structurel mais aussi fonctionnellement important.
Ce que cela signifie pour les signaux cérébraux et au‑delà
Ensemble, ces résultats montrent que TRAAK n’existe pas dans une seule forme figée. Il bascule plutôt entre états de capuchon échangé et non‑échangé dont l’abondance relative dépend de la température et des détails fins de la membrane environnante. L’état échangé est plus fréquent et devient encore plus favorisé lorsque la température augmente, suggérant que les réarrangements du capuchon peuvent contribuer à la manière dont TRAAK répond à la chaleur et aux indices mécaniques dans les cellules vivantes. En préservant les lipides natifs et en lisant précisément des conformations rares, les auteurs démontrent une méthode puissante pour relier des changements de forme microscopiques aux poches lipidiques particulières qui les accueillent. Ce cadre peut désormais être appliqué à d’autres canaux de fuite impliqués dans la douleur, l’anesthésie et les maladies neurologiques, aidant à expliquer comment de subtiles altérations de la composition membranaire et de la conformation protéique peuvent remodeler le langage électrique de nos cellules.
Citation: Ma, Y., Ackermann, K., Waheed, Q. et al. Swapped and non-swapped TRAAK states co-exist in membranes at a ratio influenced by temperature. Nat Commun 17, 3522 (2026). https://doi.org/10.1038/s41467-026-70027-9
Mots-clés: canal TRAAK, canaux de fuite du potassium, lipides membranaires, détection de la température, structure des canaux ioniques