Clear Sky Science · de
Strukturen des ZYG11B‑EloB‑EloC‑Substratkomplexes enthüllen Mechanismen des Aufbaus und der Funktion von CRL2ZYG11B
Wie Zellen ihren molekularen Abfall entsorgen
Jede Zelle in Ihrem Körper baut ständig Proteine auf und ab. Dieser beständige Turnover ist für die Gesundheit essenziell, doch die Maschinerie, die darüber entscheidet, welche Proteine bleiben oder abgebaut werden, ist außerordentlich komplex. Diese Studie beleuchtet, wie einer dieser zellulären "Türsteher", ein Protein namens ZYG11B, seine Ziele erkennt und mit Partnerproteinen zusammenarbeitet, um sie zur Zerstörung zu markieren. Die Ergebnisse erklären, wie Zellen fehlerhafte oder nicht mehr benötigte Proteine in Schach halten, und deuten auf neue Möglichkeiten hin, dieses System für Forschung und künftige Therapien zu nutzen.
Ein zelluläres Recycling‑Etikett mit besonderer Signatur
Zellen verwenden ein Anhängeschild namens Ubiquitin, um Proteine für das Recycling in einem großen, fassförmigen Kompartiment zu kennzeichnen, das sie zersetzt. Die Auswahl der zu markierenden Proteine liegt bei E3‑Ligasen, großen Komplexen, die als molekulare Kuppler fungieren. ZYG11B ist ein Baustein, der einer E3‑Ligase Zielgenauigkeit verleiht. Es erkennt Proteine, die eine sehr spezifische Signatur an ihrem N‑Ende tragen: die Aminosäure Glycin. Dieses sogenannte Gly/N‑Degron kann entstehen, wenn Proteine beim programmierten Zelltod zugeschnitten werden, wenn eine normale Lipid‑Anhängigkeitsstelle fehlschlägt oder sogar während bestimmter Virusinfektionen, wodurch ZYG11B in Prozesse wie Zellzykluskontrolle, Immunabwehr und die Art und Weise, wie einige Viren Wirtszellen kapern, eingebunden ist. 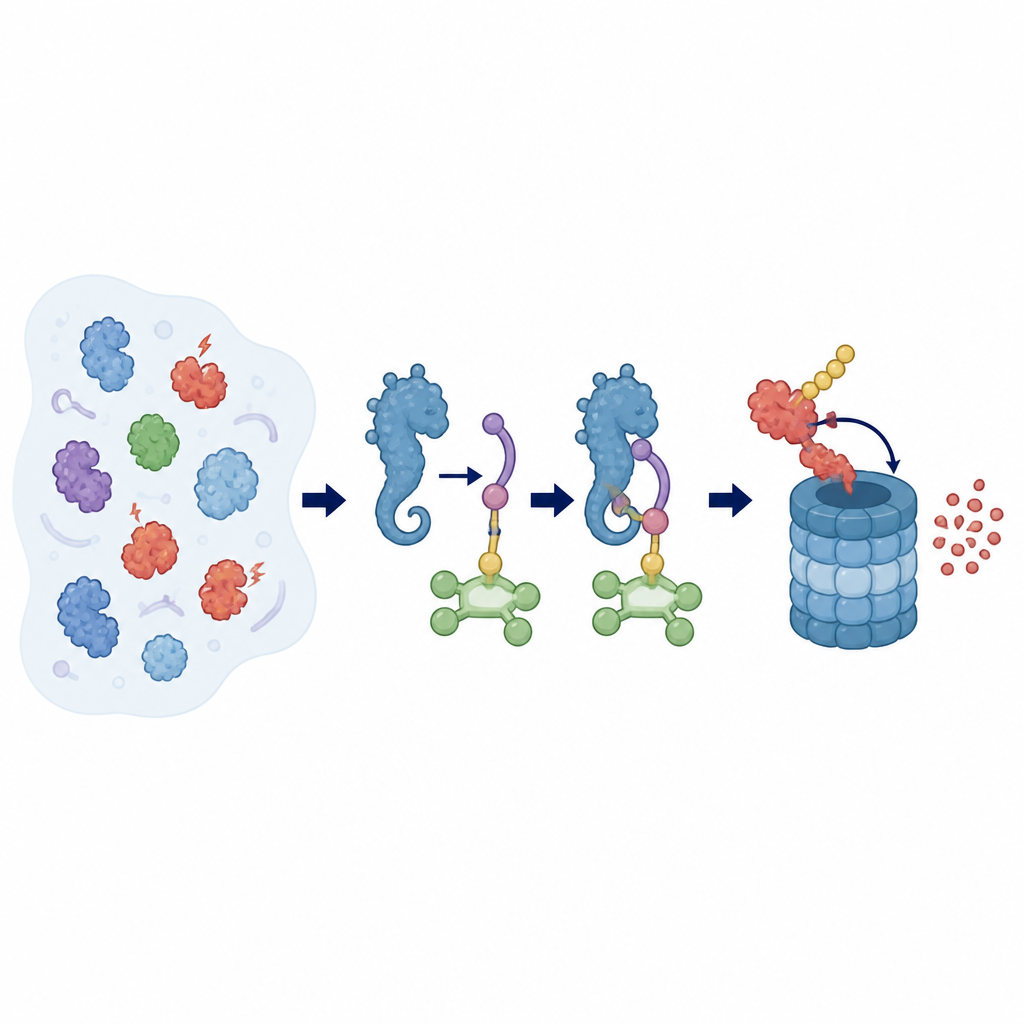
Das seepferdchenförmige Arbeitstier in Aktion sehen
Um zu verstehen, wie ZYG11B arbeitet, nutzten die Forschenden Kryo‑Elektronenmikroskopie, eine Technik, die eingefrorene Moleküle hochaufgelöst abbildet. Sie lösten die Struktur des vollständigen menschlichen ZYG11B zusammen mit zwei Partnerproteinen, EloB und EloC, und einem kurzen markierten Peptid aus einem Virusprotein. ZYG11B faltet sich zu einer auffälligen seepferdchenähnlichen Gestalt mit drei Hauptbereichen: einem Kopf, der an EloB–EloC andockt, einer zentralen Wirbelsäule aus sich wiederholenden Einheiten und einem gekrümmten Schwanz, der den Anfang des Zielproteins aufnimmt. Das glycinmarkierte Peptid liegt in einer Rinne dieses Schwanzes, wobei seine ersten drei Bausteine durch ein enges Netzwerk von Wechselwirkungen festgehalten werden, während der Rest des Peptids exponierter und flexibler bleibt.
Die Markiermaschine Stück für Stück aufbauen
Der Kopf von ZYG11B trägt ein Motiv, das fest an EloC bindet, welches wiederum EloB hält und so einen kompakten Adapter bildet. Dieser Adapter verbindet sich dann mit einem größeren Gerüstprotein namens Cul2 und einem kleinen RING‑Protein, das das ubiquitintragende Enzym anwirbt. ZYG11B nutzt drei getrennte Kontaktflächen, um EloB und EloC zu greifen, und erzeugt so eine gebogene Schleifenanordnung, die die Bindungsrinne des Schwanzes nahe an den katalytischen Teil der Ligase bringt. Als das Team Schlüsselkontakte im Kopf von ZYG11B oder in der Rinne, die das glycinmarkierte Peptid hält, veränderte, konnten Zellen Proteine mit diesem Tag nicht mehr effizient abbauen. Das zeigte, dass sowohl Substratbindung als auch Adapterdocking für die Qualitätskontrolle dieses Systems unerlässlich sind.
Wenn eins zu zwei wird: Die Kraft des Paarens
Eine unerwartete Wendung war, dass ZYG11B nicht immer allein arbeitet. Die Strukturdaten zeigten, dass sich zwei ZYG11B‑Moleküle Rücken an Rücken paaren können, jeweils mit eigenem Adapter und markiertem Peptid, und so ein symmetrisches Dimer bilden. Dieses Paaren umfasst alle drei Regionen von ZYG11B und begräbt eine große Kontaktfläche, wodurch eine stabile Einheit mit zwei aktiven Bindungsrinnen entsteht, die in entgegengesetzte Richtungen weisen. Die Forschenden erstellten Modelle der vollständigen Ligase in dieser paarigen Form und zeigten, dass sie weiterhin die katalytischen Komponenten für die Markierung aufnehmen kann. In Reaktionen im Reagenzglas und zellbasierten Tests zeigte eine Mutante von ZYG11B, die die Dimerbildung stört, deutlich schwächere Markierung und Abbau der Zielproteine, was darauf hindeutet, dass der gepaarte Zustand die Effizienz des Systems steigert. 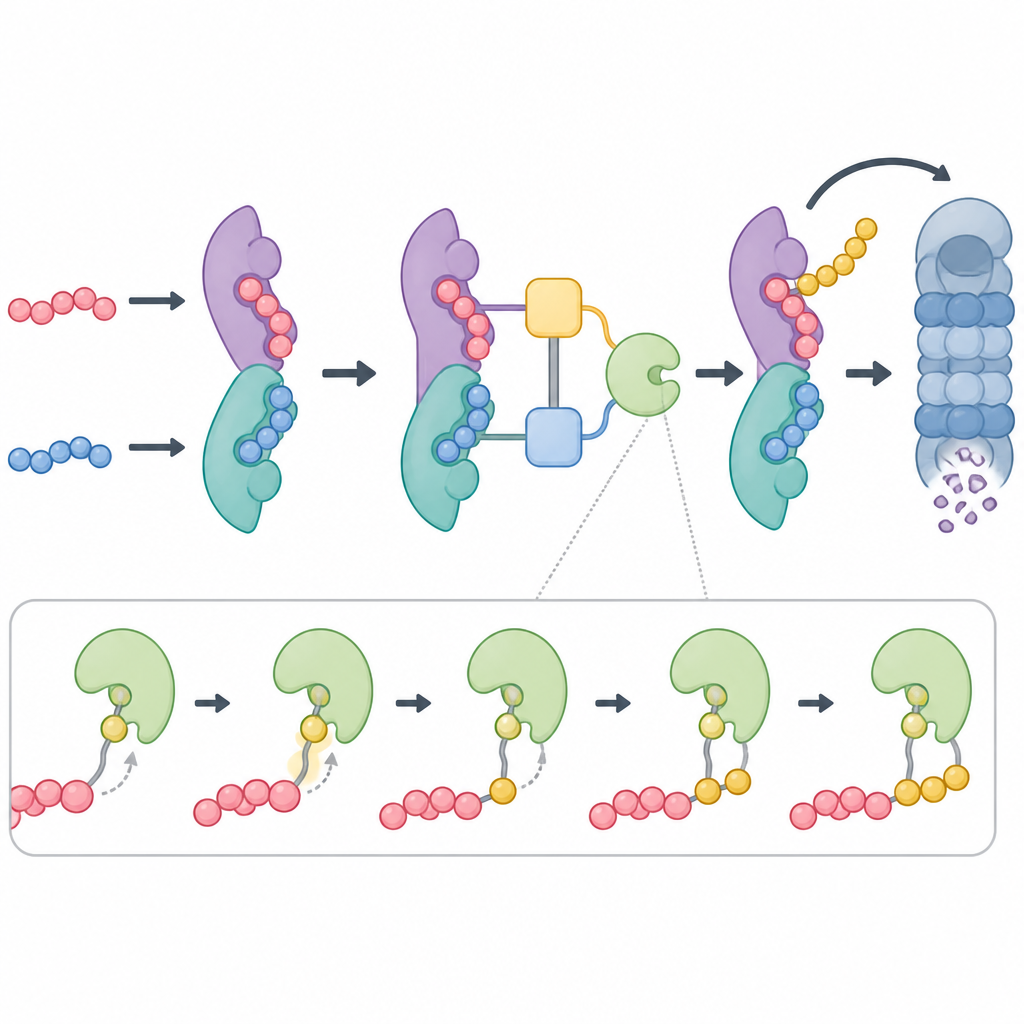
Warum diese Erkenntnisse für Gesundheit und Design wichtig sind
Zusammen legen die Ergebnisse nahe, dass die ZYG11B‑basierte Ligase zwischen Einzel‑ und Doppelzuständen wechseln kann, wobei beide vermutlich in Zellen vorkommen, der gepaarte Zustand jedoch eine führende Rolle bei effizienter Markierung und Zerstörung spielt. Indem diese Arbeit die detaillierte Gestalt von ZYG11B und genau zeigt, wie es sowohl seine Partner als auch seine Ziele greift, liefert sie eine Blaupause für das Design von kleinen Molekülen oder maßgeschneiderten Degradatoren, die ZYG11B an neue Ziele rekrutieren. Langfristig könnten solche Werkzeuge es Wissenschaftlern ermöglichen, gezielt schädliche Proteine zu entfernen, was ein kraftvolles Mittel wäre, zelluläre Signalwege zu untersuchen und potenziell Krankheiten anzugehen, die mit der Proteinqualitätskontrolle zusammenhängen.
Zitation: Lin, N., Feng, H., Geng, Y. et al. Structures of ZYG11B-EloB-EloC-substrate complex reveal mechanisms of CRL2ZYG11B assembly and function. Nat Commun 17, 4648 (2026). https://doi.org/10.1038/s41467-026-71318-x
Schlüsselwörter: Proteinabbau, Ubiquitin‑Ligase, ZYG11B, Gly N Degron, Cryo‑Elektronenmikroskopie