Clear Sky Science · ar
مهيّئ بروتيني يوسّط السيطرة المعتمدة على Ap4A على أسيتلة البروتينات
لماذا تهم هذه الإشارة الصغيرة للإجهاد
داخل كل خلية، تُشغّل البروتينات أو تُعطّل باستمرار عبر وسوم كيميائية صغيرة. أحد هذه الوسوم، المُسمّى الأسيتلة، يعيد تشكيل كيفية استخدام الخلايا للطاقة ونسخ الحمض النووي وبناء مكوّنات جديدة. تكشف هذه الورقة كيف يساعد جزيء إنذار صغير، Ap4A، البكتيريا على إعادة ضبط وسم البروتين هذا بسرعة أثناء الإجهاد. ومع أن العمل أُجري في بكتيريا تربوية، فإن اللاعبين الأساسيين يشبهون ما يوجد في خلايانا، ما يشير إلى طرق مشتركة واسعة لدى الكائنات الحية للاستجابة لتغير الظروف.
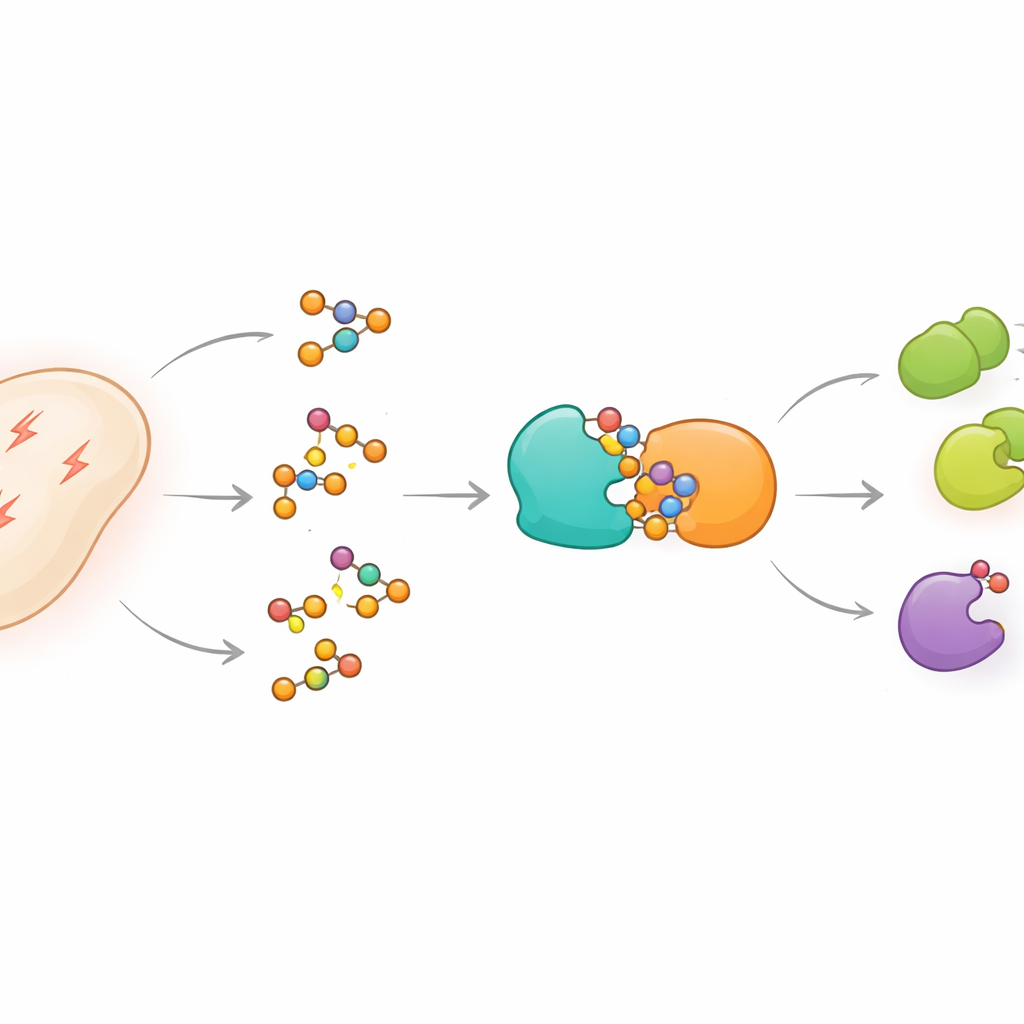
مفتاح تشغيل/إيقاف كيميائي للبروتينات
تحمل العديد من البروتينات «مقابض» حيث يمكن للخلايا إضافة مجموعة أسيتيل أو إزالتها، مما يغيّر سلوك البروتين بشكل دقيق دون إعادة بنائه من الصفر. تضيف إنزيمات متخصصة الوسم، بينما تزيله أخرى للحفاظ على توازن النظام. في البكتيريا Bacillus subtilis، هناك إنزيم رئيسي يُدعى AcsA يصنع الأسيتيل‑CoA، الجزيء المركزي للوقود واللبنة البنائية. يُثبط نشاط AcsA عندما يُأسَتَل ويعود للعمل عندما يزيل إنزيم شريك، الديأسيتيلارز AcuC، مجموعة الأسيتيل. تقع الجينات المسؤولة عن آلية أسيتلة AcsA بجانب جين ثالث غامض، acuB، ما يوحي بأنه يلعب دورًا ذا صلة لكن لم يُكشف من قبل.
كشف المكبح الخفي ضد إزالة الأسيتلة
سعى المؤلفون لاكتشاف وظيفة AcuB. عبر استخلاص AcuB من خلايا حية وتحديد البروتينات التي جاءت معه، وجدوا أن AcuB يُشكّل معقدًا ثابتًا مع AcuC، الديأسيتيلارز. أظهرت تجارب أنبوب الاختبار بالبروتينات المنقاة أنه عندما يتواجد AcuB، لم يعد AcuC قادرًا بكفاءة على إزالة مجموعات الأسيتيل من أهدافه، بما في ذلك AcsA وبروتينات أخرى مشاركة في تصنيع البروتين، وبناء جدار الخلية، والتحكم في نسخ الحمض النووي. جوهريًا، يعمل AcuB كمكبح مادي على AcuC، يحافظ على العديد من البروتينات في حالتها المسيتلة والمعدّلة.
كيف يُثبّت جزيء الإنذار Ap4A المكبح
ثم يربط البحث هذا المكبح بنظام إنذار أوسع للإجهاد. في ظروف قاسية، تتراكم البكتيريا Ap4A، وهو جزيء صغير طالما أُشتبه في كونه إشارة خطر. باستخدام تجارب الارتباط والعمل البنيوي عالي الدقة، يظهر الباحثون أن Ap4A يناسب بإحكام وحدتين حسّاسيتين مزدوجتين في AcuB، مما يثبّت بروتين AcuB بشكل كبير. عندما يرتبط Ap4A، يصبح AcuB أكثر مقاومة للحرارة كما يرتبط بقوة أكبر مع AcuC. تشير البيانات البنيوية والمحاكاة إلى أن ذراعًا في AcuB تتحرك لتسد فم الموقع الفعّال في AcuC، مانعة وصول ذيول البروتين المسيتلة. بوجود Ap4A، يصبح هذا السداد أوضح، مما يُضعف قدرة AcuC على نزع وسوم الأسيتيل بشكل أكبر.
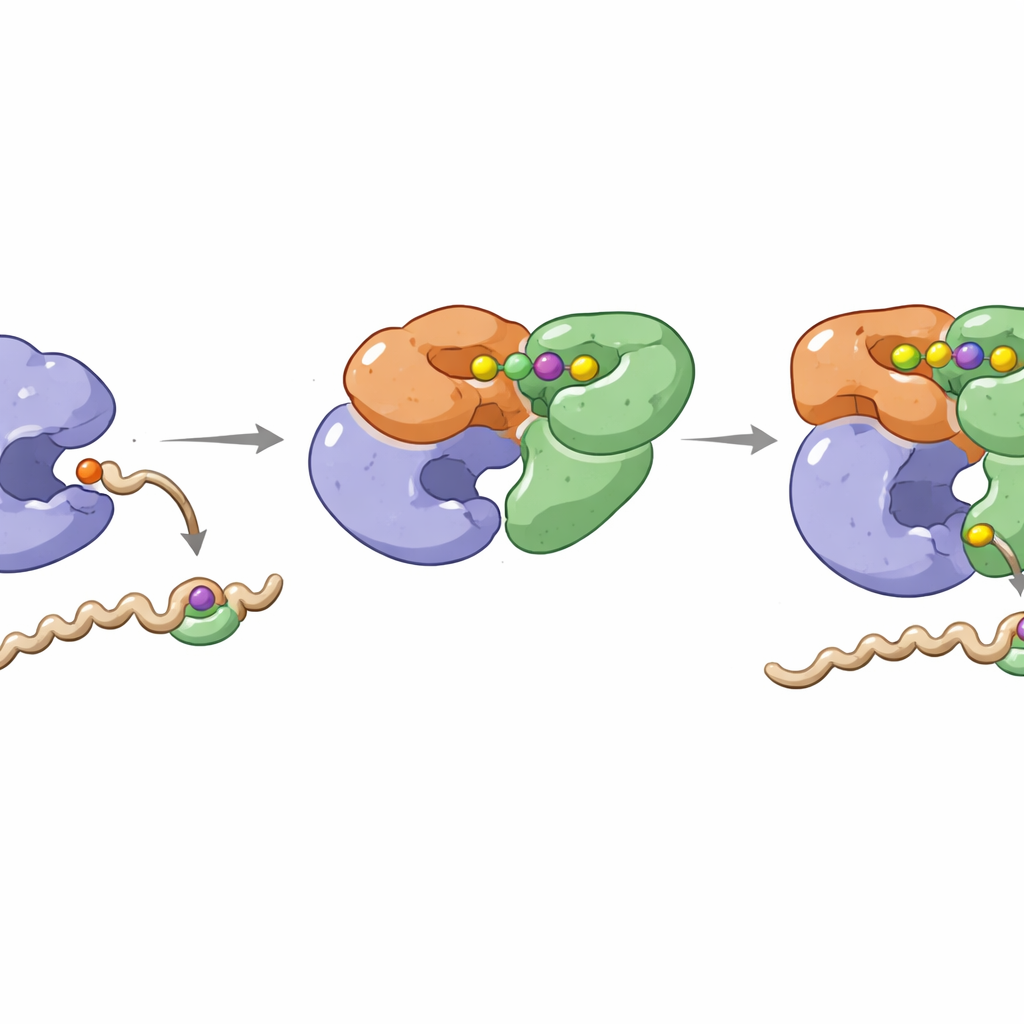
شبكة خاضعة للسيطرة بالإجهاد تمس وظائف خلوية عديدة
بما أن AcuC يمكن أن يؤثر على عدة بروتينات غير مترابطة، فإن هذه الوحدة التنظيمية الواحدة لها امتداد واسع. عندما تكون مستويات Ap4A منخفضة، يكون AcuB غير مستقر نسبيًا، ويكون AcuC حرًا في إزالة مجموعات الأسيتيل في أنحاء الخلية، مما يعزز نشاط الإنزيمات وإنتاج الأسيتيل‑CoA القوي. عندما يرفع الإجهاد مستويات Ap4A، يتراكم AcuB المرتبط بـAp4A ويشدّد قبضته على AcuC. نتيجة لذلك، تبقى بروتينات متعددة مسيتلة: يقل نشاط AcsA في تصنيع الأسيتيل‑CoA، وتبقى عوامل المشاركة في الترجمة وبناء جدار الخلية ونسخ الحمض النووي في حالات معدّلة. يقترح المؤلفون أن هذا يمنح الخلية وسيلة سريعة لتنسيق استخدام الطاقة والعمليات الأساسية مع ظروف الإجهاد، دون الحاجة أولًا لتشغيل أو إيقاف الجينات.
ما المغزى خارج هذه البكتيريا
تكشف هذه الدراسة عن AcuB كمُهيّئ يحول إشارة إنذار عامة، Ap4A، إلى تحكّم موجه في أسيتلة البروتين عن طريق تثبيط إنزيم شبيه بـHDAC. نظرًا لأن وحدات حسّية مماثلة وأنظمة أسيتلة وأقارب HDAC توجد من البكتيريا إلى الإنسان، قد يتردد صدى الآلية الموصوفة هنا في خلايا أكثر تعقيدًا حيث تلعب HDACs دورًا محوريًا في التحكم الجيني وتُعد أهدافًا دوائية رئيسية. ببساطة، تبيّن الدراسة كيف يمكن لجزيء إجهاد صغير أن يثبّت مكبحًا بروتينيًا يثبط إنزيمًا رئيسيًا، فيغيّر العديد من مفاتيح الخلية دفعة واحدة لمساعدة الخلية على التكيّف مع الشدائد.
الاستشهاد: Zheng, L., Young, M.K.M., Steinchen, W. et al. A protein adaptor mediating Ap4A-dependent control of protein acetylation. Nat Commun 17, 3089 (2026). https://doi.org/10.1038/s41467-026-70006-0
الكلمات المفتاحية: أسيتلة البروتين, استجابة بكتيرية للإجهاد, جزيء الإنذار Ap4A, تنظيم دياستيلاز الهيستون, استقلاب الأسيتيل‑CoA