Clear Sky Science · en
The local mechanostructural properties of protein cargoes regulate nucleocytoplasmic transport
How protein shape steers traffic into the cell nucleus
Every second, thousands of proteins must pass in and out of the cell’s control center, the nucleus. This traffic is tightly regulated, because it decides which genes are turned on and how cells respond to their environment. This study reveals that not only the identity of a protein, but also which end of it goes first and how floppy or stiff that end is, can strongly influence how quickly it crosses the nuclear border.
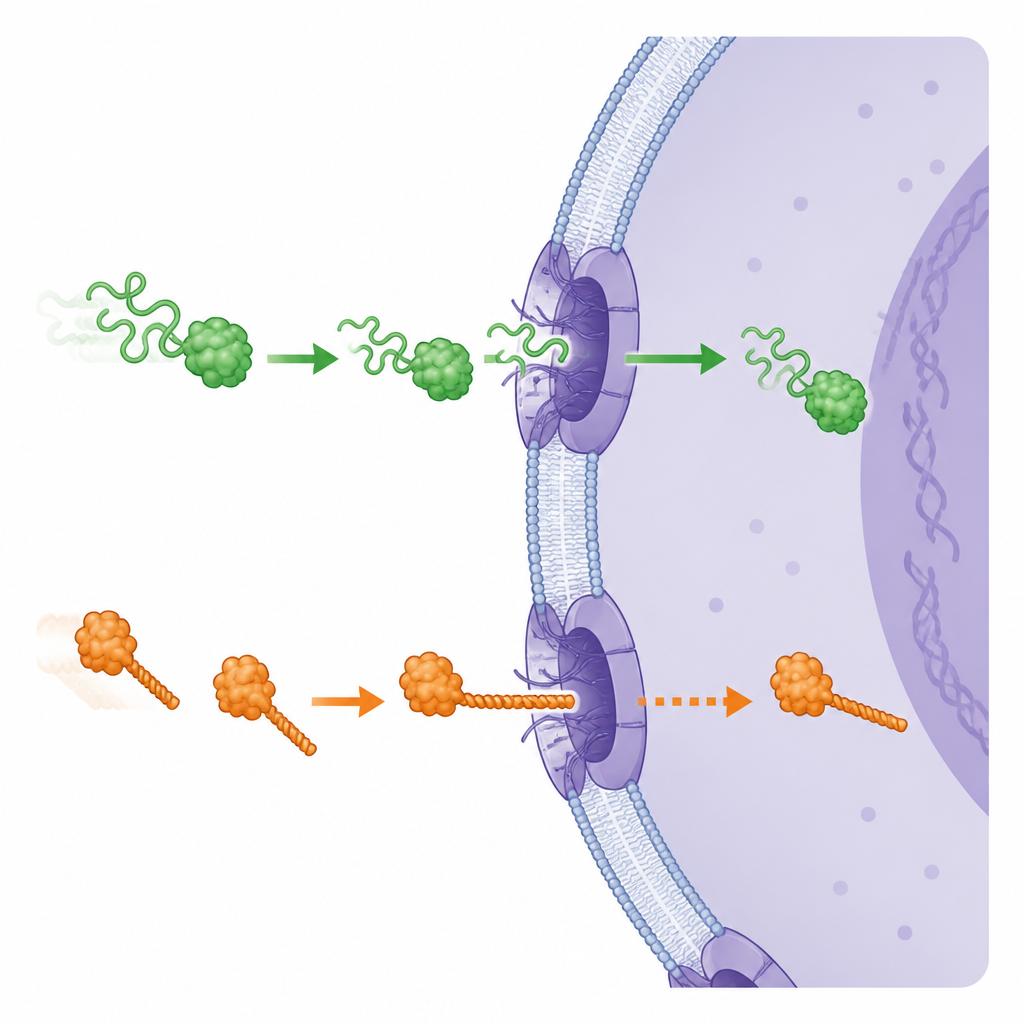
The nuclear doorway as a selective gateway
Proteins move between the cytoplasm and the nucleus through large channels called nuclear pore complexes. These pores are unlike many other cellular gates that rely on energy-driven motors to pull proteins through tight openings. Earlier work showed that proteins that are easier to mechanically unfold tend to enter the nucleus faster. The authors asked whether, even for a single protein, local softness or stiffness near one end could make a difference to how it threads through the pore and how fast it travels.
Pulling on proteins one by one
To answer this, the researchers combined three powerful approaches. First, they used magnetic tweezers to grab individual protein molecules and gently pull them apart, revealing how much force is needed to unfold different regions. Second, they used computer simulations to visualize how each end of a protein starts to unravel when force is applied from a specific direction. Third, they engineered light-controlled protein constructs in living cells, so that a brief flash of blue light could trigger proteins to move into or out of the nucleus while microscopes tracked their journey in real time.
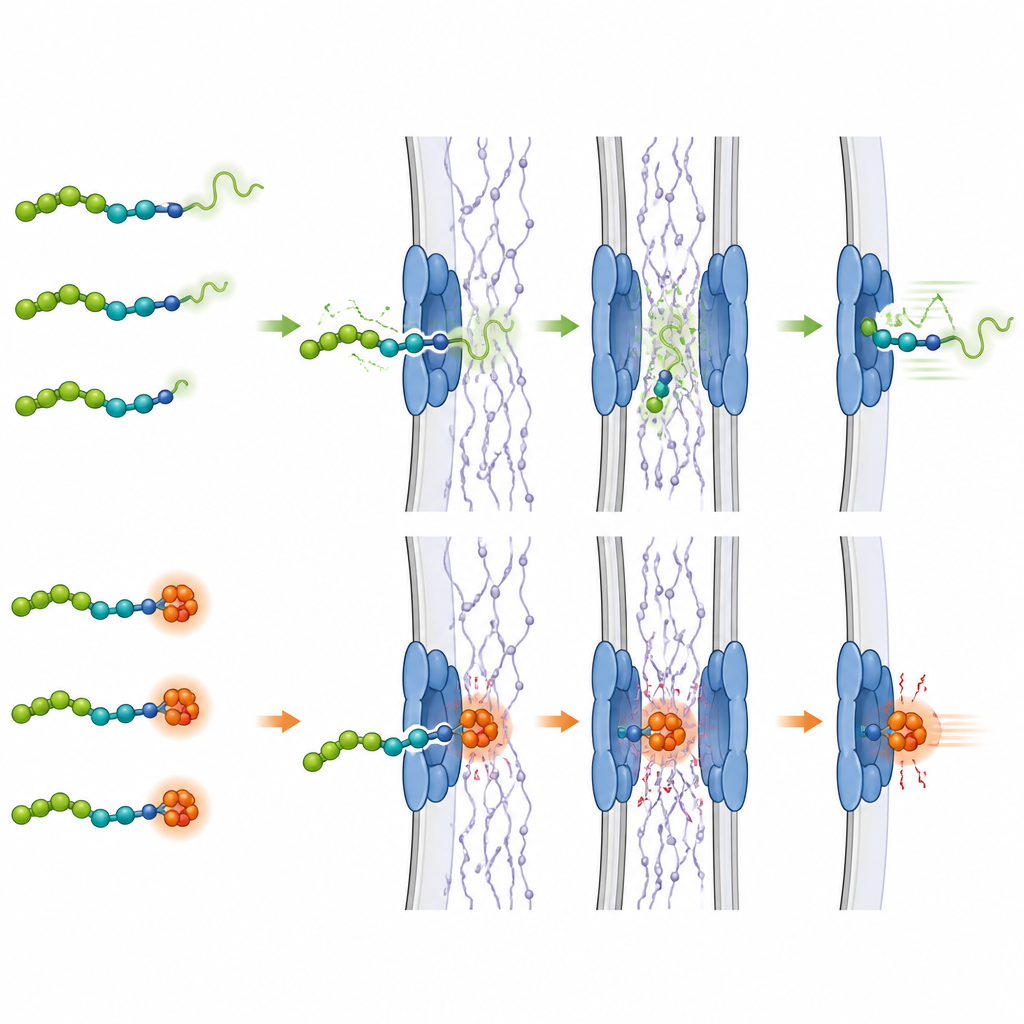
Soft ends go first and move faster
Across several very different proteins, a clear pattern emerged. When a protein was arranged so that its more flexible, mechanically weaker end approached the pore first, it entered the nucleus faster and accumulated there to higher levels than when the stiffer end led the way. For example, a fluorescent protein called mCherry unfolds more easily at its N-terminal end, and it showed quicker nuclear import when that end entered first. When the team rewired the protein so that its vulnerable segment was relocated, both its unfolding behavior and its import speed changed in lockstep. Similar orientation effects were seen for other test proteins with varied shapes and stabilities, and even for a bacterial protein whose ends are normally locked together by special internal bonds. Breaking just one of these bonds made that end unfoldable and, when placed at the front, sped up its passage through the pore.
Export, nuclear helpers, and real gene regulators
The same idea also applied, though more modestly, to proteins leaving the nucleus. Export was generally faster when the protein exited with its more pliable end first, suggesting that local softness matters for both directions of travel. The researchers further found that a component of the nuclear pore, called Nup153, is particularly important for sensing these floppy regions: when Nup153 was reduced in cells, the difference between soft-end-first and stiff-end-first transport largely disappeared. To see whether this principle matters for natural gene regulators, the team examined SMAD4, a protein that controls cell growth signals. They showed that SMAD4 enters the nucleus more rapidly when it is oriented so that its naturally softer end, which also carries its built-in nuclear address tag, goes in first.
Hints from the wider family of gene switches
Looking beyond individual examples, the authors turned to bioinformatics to survey more than a thousand human transcription factors, the proteins that switch genes on and off. They predicted where nuclear localization signals, the short address tags that tell transport machinery where to bind, sit along each protein sequence and how structurally ordered or disordered the surrounding regions are. Strikingly, in many transcription factors the stretch from the nuclear localization signal to the nearest end of the protein is more disordered, and therefore likely more flexible, than the average for that protein. This suggests that evolution may have favored designs where the nuclear address tag is embedded in a floppy segment that can more easily begin to unfold and engage with the pore.
Why this matters for cellular control
Together, these findings show that the local mechanical properties of proteins add an extra layer of control to nuclear traffic. Proteins whose leading end is softer and more disordered can begin to unravel slightly, revealing sticky patches that interact with flexible strands inside the nuclear pore, helping them move more quickly. By reshaping or mutating specific regions, it may be possible to tune how efficiently proteins reach the nucleus. This work links the nanoscale feel of a protein’s backbone to the large-scale regulation of gene activity, offering new ways to think about designing protein-based therapies and tools that must reliably find their way into the cell’s command center.
Citation: Tapia-Rojo, R., Milmoe, N., Paracuellos, P. et al. The local mechanostructural properties of protein cargoes regulate nucleocytoplasmic transport. Nat. Phys. 22, 770–783 (2026). https://doi.org/10.1038/s41567-026-03242-2
Keywords: nuclear pore complex, protein mechanics, nucleocytoplasmic transport, transcription factors, protein unfolding