Clear Sky Science · en
The nitrogen starvation-induced inhibitor Rts3 restrains Sit4/PP6 to gate quiescence downstream of TORC1
How Cells Decide to Rest
Our bodies, and even simple yeast cells, must sometimes switch from busy growth to a low‑power resting mode to survive hard times. This paper explores how baker’s yeast senses a shortage of nitrogen, a key nutrient, and chooses not only to pause growth but also how deeply to go into this resting state. The authors uncover a tiny protein, Rts3, that works like a molecular brake, helping cells enter a protective pause without getting so deeply shut down that they cannot easily wake up again when food returns.
The Main Growth Switch in Cells
At the heart of this story is a protein control center called TORC1, which acts as a growth switch in organisms from yeast to humans. When nutrients are plentiful, TORC1 encourages cells to build proteins, divide, and store energy, while suppressing recycling and clean‑up processes. When nutrients run low, TORC1 activity falls, and cells turn on survival programs instead. In yeast, low TORC1 activity helps trigger a state called quiescence: metabolism slows, stress resistance rises, and specific genes turn on to get the cell through famine. Problems with TORC1 control in humans are linked to cancer, obesity, and brain disorders, so understanding how this switch is tuned, even in yeast, has broad importance.
Finding a Hidden Brake Protein
The researchers set out to map the landscape of enzymes that remove phosphate tags from proteins after TORC1 is shut down with the drug rapamycin. Using chemical “bait” beads and mass spectrometry, they fished out many phosphatases and their partners. Among these, one little‑known protein, Rts3, stood out as strongly enriched when TORC1 was inhibited. Earlier hints suggested that Rts3 might partner with a group of phosphatases related to the human PP6 enzyme, especially one called Sit4 in yeast. By tracking tagged Rts3, the team showed that Rts3 levels are very low during normal growth but rise quickly during nitrogen starvation or rapamycin treatment, then fall again within minutes when amino acids are added back.
A Molecular Brake on a Key Enzyme
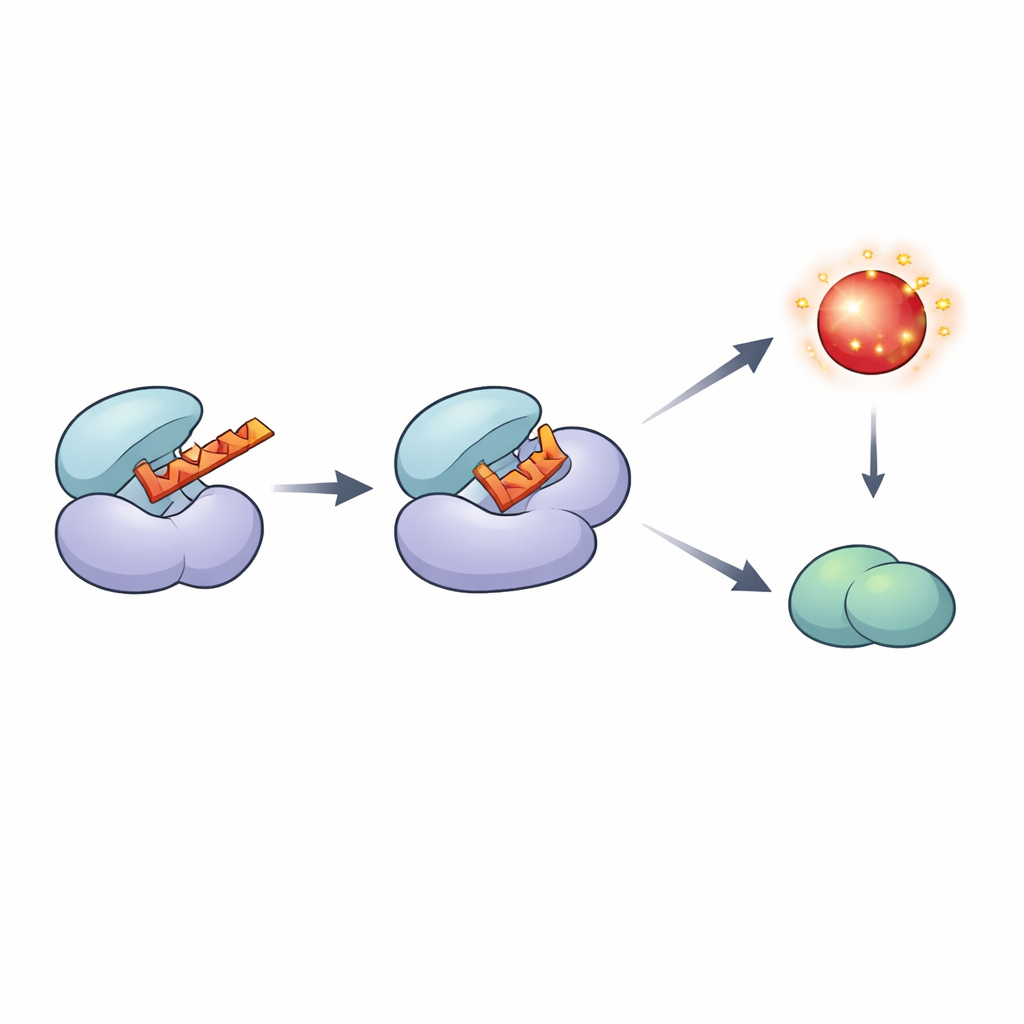
Digging deeper, the authors discovered that Rts3 is not a standard helper subunit for broad‑acting phosphatases, as once thought. Instead, it directly binds Sit4 when Sit4 is paired with two specific partners, Sap185 or Sap190. Structural predictions and computer simulations revealed that a short alpha‑helix segment of Rts3 fits snugly into the active groove of Sit4, physically blocking the site where substrates would normally bind. In test‑tube assays, adding Rts3 sharply reduced Sit4’s activity at very low concentrations, while mutant versions of Rts3 missing key contact points could no longer inhibit Sit4 or protect cells from rapamycin. Inside cells, this means Rts3 selectively reins in one branch of the Sit4 network rather than shutting the enzyme down everywhere.
Balancing Survival Programs During Starvation
When nitrogen runs low and TORC1 is turned down, Sit4‑Sap185/190 helps activate a range of survival programs. These include genes controlled by Gln3 and Gat1 that let cells use poor nitrogen sources, genes regulated by Rtg3 that adjust mitochondrial metabolism, and pathways that slow protein production through stress‑responsive factors. Using large‑scale phosphoproteomics and protein profiling, the authors showed that boosting Rts3 dampens many of these starvation responses, while removing Rts3 lets them run too strongly. Rts3 also associates with ribosomes and affects factors that decide whether ribosomes keep making proteins or enter a “hibernating” state. Yeast lacking Rts3 grow slightly better on poor nitrogen but pay a price: they survive less well during long‑term starvation, suggesting that unrestrained Sit4 pushes cells into an unsafely deep quiescent state.
A Fast Feedback Loop for Reversible Rest
Strikingly, the same transcription factor Gln3 that Sit4 activates during starvation also turns on the RTS3 gene, creating a feedback loop. As Rts3 builds up, it feeds back to limit Sit4 signaling and prevent overshooting. When nutrients return, TORC1 is reactivated and directly tags Rts3 at a specific site, which allows a ubiquitin‑based protein‑destruction machine to rapidly remove Rts3. This two‑part control—gene activation by nitrogen‑stress signals and swift protein destruction when food comes back—gives cells a snap‑like switch: they can deepen their resting state when needed but also shed the brake quickly to resume growth.
Why This Matters Beyond Yeast
In simple terms, this study shows how a cell installs a small, removable brake pedal on a major growth‑control pathway. Rts3 helps yeast strike a sweet spot between protection and flexibility: enough slowdown to endure famine, but not so much that recovery is clumsy or delayed. Because the Sit4 enzyme is closely related to human PP6, which has been linked to cancers and therapy resistance, similar helix‑based inhibitors may tune quiescence and stress responses in our own cells. Understanding this fine‑tuned braking system could eventually inform ways to better control cell dormancy in disease or aging.
Citation: Dokládal, L., Péli-Gulli, MP., Alba, J. et al. The nitrogen starvation-induced inhibitor Rts3 restrains Sit4/PP6 to gate quiescence downstream of TORC1. Nat Commun 17, 3256 (2026). https://doi.org/10.1038/s41467-026-69693-6
Keywords: cellular quiescence, TORC1 signaling, protein phosphatase Sit4/PP6, nitrogen starvation, yeast stress response