Clear Sky Science · en
Organelle contact sites in cancer cells
How tiny bridges inside cells could change cancer treatment
Inside every cell, miniature compartments called organelles carry out different jobs, from making energy to recycling waste. This review explains how these structures do not work in isolation but constantly touch and talk to each other through tiny bridges. Because these contact sites help control how cancer cells grow, move, and resist treatment, understanding them could open up new ways to diagnose and treat tumours.
Cellular neighbourhoods and their hidden conversations
Organelles such as mitochondria, the endoplasmic reticulum, lysosomes, lipid droplets, peroxisomes, the nucleus, and the cell surface form close, long-lasting contacts without actually fusing. At these junctions, they swap ions like calcium, share fats and cholesterol, and pass signals that fine-tune metabolism, stress responses, and cell survival. The review describes how these contact sites are flexible structures that remodel when cells face stress or changing fuel demands, acting as central hubs for keeping the internal environment in balance.
Energy factories and the stress network
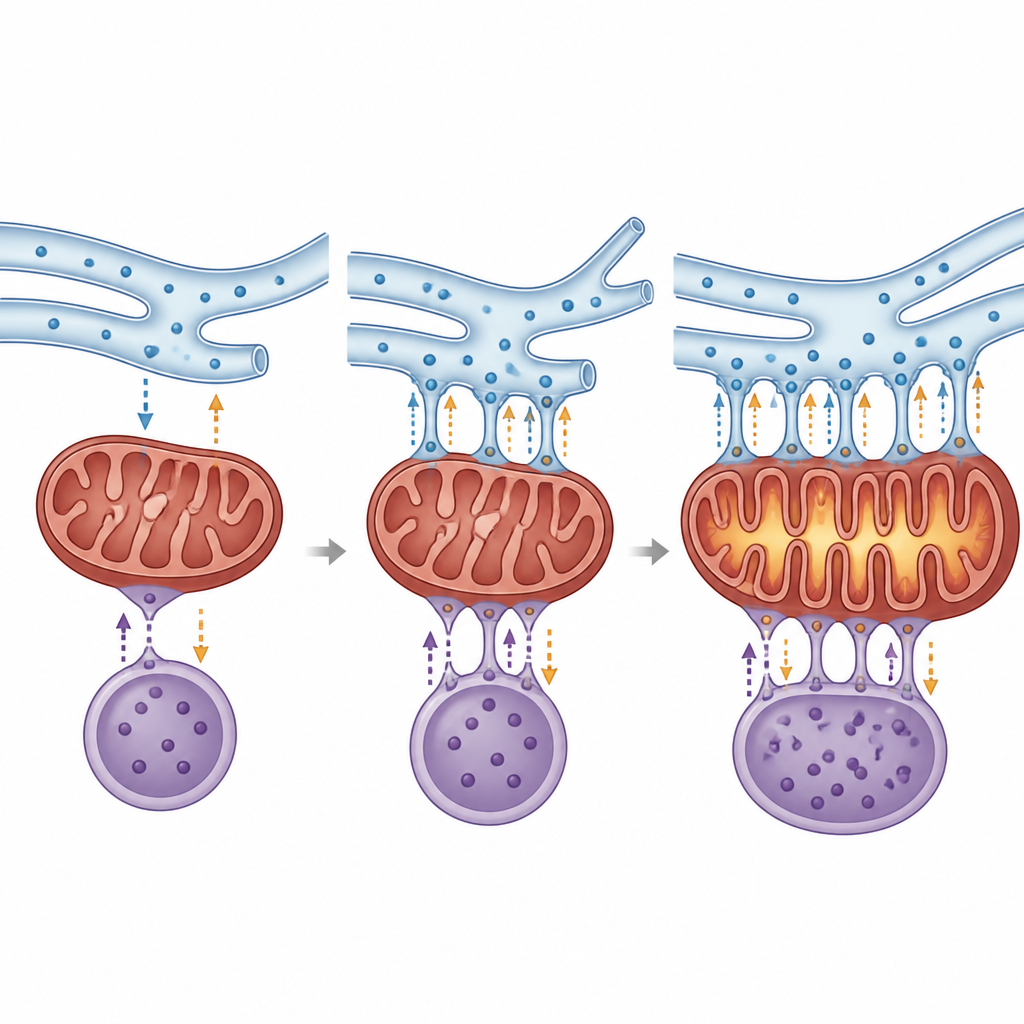
A major focus is the partnership between mitochondria, the cell’s power plants, and the endoplasmic reticulum, which helps fold proteins and store calcium. Their contact regions control energy production, redox balance, and sensitivity to cell death. In cancer cells, many of the tethering proteins that hold these structures together are altered. Some changes increase calcium flow into mitochondria, boosting fuel burning and growth, while others shield the cell from the calcium overload that would normally trigger its self-destruction. Proteins such as FUNDC1, PERK, mitofusins, and Sigma-1 receptor can either support or restrain tumour growth depending on the tissue and context, highlighting how the same physical bridge can play opposite roles in different cancers.
Waste stations, fat stores, and iron traffic
Contacts between the endoplasmic reticulum and late endosomes or lysosomes help position these recycling stations in the cell and govern how cholesterol and other lipids move. In many tumours, tethers like STARD3, VAP proteins, NPC1, ORP5, and Protrudin are hijacked to boost cholesterol supply, support hormone production, or drive the formation of invasive protrusions that cut through surrounding tissue. Other contact sites link mitochondria with lysosomes and control the turnover of damaged mitochondria, iron distribution, and sensitivity to a type of iron-dependent cell death called ferroptosis. Proteins such as DMT1, TRPML1, Rab7, and BDH2 shape how iron and calcium are shuttled between these compartments, with direct consequences for how easily cancer cells invade, survive low oxygen, or respond to therapy.
Fat depots, peroxisomes, and the cell surface
The review also outlines how lipid droplets, which store fats, plug into mitochondria and the endoplasmic reticulum to feed energy-hungry cancer cells. Tethers involving PLIN proteins, CPT1A, FATP4, MIGA, and VPS13 family members channel fatty acids from droplets into mitochondria for burning or building new membranes. Peroxisomes, which help process fats and hydrogen peroxide, are wired to the endoplasmic reticulum through ACBD5 and VAP proteins, coordinating fatty acid breakdown and membrane building. At the cell surface, junctions between the endoplasmic reticulum and the plasma membrane, organised by STIM, ORAI, extended synaptotagmins, ORPs, PTP1B, and GRAMD proteins, manage calcium entry and lipid flow that in turn shape cell movement, blood vessel growth, and resistance to cell death.
Crossroads for immunity and multi-organellar hubs
Because many immune sensors sit at or near these contact sites, the same bridges that fuel cancer cells also influence how the immune system recognises and attacks tumours. Changes in endoplasmic reticulum–mitochondria or endoplasmic reticulum–endosome links can tune inflammasome activity, the cGAS–STING pathway, and forms of cell death that alert immune cells. The authors describe higher-order junctions where three or more organelles meet, such as endoplasmic reticulum–mitochondria–lysosome or endoplasmic reticulum–mitochondria–lipid droplet clusters. These multiway hubs reshape the local protein makeup, coordinate energy use, and help generate inflammatory lipids, further tying metabolism to immune control.
Drugs that target the cell’s internal bridges
Because many tethers use enzymes or chaperones already studied in cancer, researchers have begun testing compounds that alter organelle contacts. Some molecules act directly on the bridges, such as agents that disrupt the GRP75-based link between the endoplasmic reticulum and mitochondria or block the cholesterol shuttle STARD3, tipping cancer cells toward energy crisis and death. Others change calcium handling or lysosome function and only indirectly remodel contact sites. While most work is still at the cell or animal stage, these efforts suggest that fine-tuning the distance and dialogue between organelles could complement existing treatments, for example by making tumours more sensitive to chemotherapy or immune attack.
Why these tiny junctions matter for future cancer care
The article concludes that organelle contact sites act as master switches for calcium, fat handling, metabolism, stress responses, and immune signalling, all of which define how dangerous a tumour becomes and how well it responds to drugs. Although much remains to be learned, especially about how these bridges behave in different cancer types and patients, the authors argue that mapping and pharmacologically adjusting these microscopic junctions may offer a new layer of precision in cancer therapy beyond genes and single proteins.
Citation: Celotti, I., Scavezzon, M., Toffanin, S. et al. Organelle contact sites in cancer cells. Cell Death Dis 17, 454 (2026). https://doi.org/10.1038/s41419-026-08674-5
Keywords: organelle contact sites, cancer metabolism, mitochondria endoplasmic reticulum, calcium signalling, lipid trafficking