Clear Sky Science · zh
PFKM 控制骨骼肌分化过程中的代谢转向
为什么肌细胞需要聪明地利用糖
当我们运动或只是从椅子上站起来时,骨骼肌会立即启动,燃烧糖以驱动每一次收缩。但年轻的肌肉干细胞与成熟的肌纤维并不以相同方式处理糖。本研究揭示了一个内在的切换系统——以单一酶 PFKM 为核心——决定葡萄糖是被用于立即供能,还是被改道以保护和重建细胞。理解这一开关可能为治疗肌无力、肌肉衰老和遗传代谢疾病开辟新途径。
细胞燃料的红绿灯
进入细胞的葡萄糖可以走几条不同路线。一条主要道路是糖酵解,它将糖迅速转化为能量和用于生长肌纤维的构件。另一条路线是戊糖磷酸途径,它产生可抵御氧化应激并支持 DNA 与脂质合成的分子。研究者将注意力集中在位于糖酵解关键交汇处的磷酸果糖激酶‑1 的肌肉亚型 PFKM。通过在激活称为 Wnt 的生长信号后的数分钟到数小时内测量数百种代谢物,他们发现早期糖酵解中间产物迅速堆积,而 PFKM 的产物下降,同时戊糖磷酸途径的代谢物上升。这表明细胞主动抑制 PFKM,将葡萄糖改道到保护性化学反应而非单纯的能量生产。 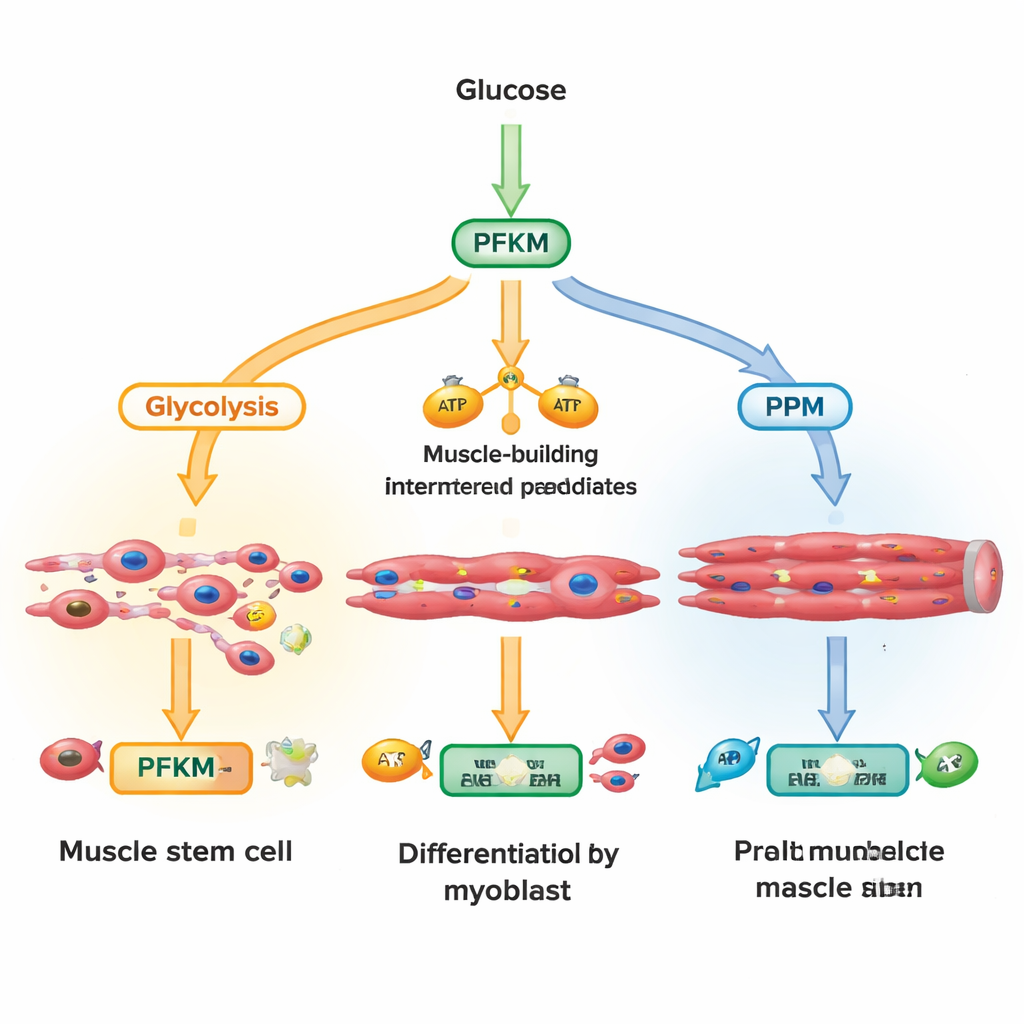
标记并移除门控酶
为弄清 PFKM 如何被调控,团队追踪了该蛋白在细胞内的位置。在静息条件下,PFKM 在细胞质中分散分布。Wnt 刺激后数分钟内,它聚集成明亮的点,且与溶酶体——细胞的蛋白降解隔间——重叠。两个同源酶 PFKL 和 PFKP 未见移动或水平变化,表明只有肌肉型 PFKM 被定向降解。生化实验显示,阻断溶酶体功能可阻止 PFKM 的丧失,而抑制蛋白酶体(另一种主要的蛋白处置系统)则无效。序列分析发现 PFKM 含有一段独特的“甲基精氨酸降解子(methyl arginine degron)”基序。酶 PRMT1 在该基序的一个精氨酸上添加特定的甲基标记,该标记使微自噬装置(包括蛋白 VPS4)能够将 PFKM 拉入溶酶体进行降解。禁用 PRMT1 或 VPS4 会稳定 PFKM 并阻止其去除。
从干细胞到能工作的肌纤维
利用大型人类单细胞数据集,作者绘制了多种肌细胞类型中的 PFKM 水平。处于休眠状态、直到需要修复时才被动员的肌肉干细胞几乎不含 PFKM,但表达大量戊糖磷酸途径基因和溶酶体相关组分。随着细胞向肌肉命运确认并融合成多核纤维,PFKM 的转录本和蛋白水平急剧上升,而 Wnt 靶基因和溶酶体基因则下降。在培养的人和小鼠肌细胞中,Wnt 在早期单核细胞中迅速将 PFKM 驱入溶酶体,但在成熟的多核纤维中则不见此现象。该模式支持这样一个模型:未分化细胞通过溶酶体降解维持低 PFKM,以偏向保护性代谢;随后在向高能耗收缩纤维转变时重新表达 PFKM。
当开关卡住会发生什么
为测试 PFKM 对肌肉构建的重要性,团队通过 RNA 干扰降低其水平。PFKM 低表达的细胞难以开启典型的肌肉基因,产生更少的肌球蛋白蛋白,形成的多核纤维更少且更小,尽管总体细胞数并未改变。代谢物谱分析表明下游糖酵解中间产物和三羧酸循环燃料减少,但戊糖磷酸途径基因和标志物增加,同时对氧化应激的抵抗增强。重要的是,向细胞补充 3‑磷酸甘油酸(一种通常位于 PFKM 下游的糖酵解中间产物)可挽救许多分化缺陷。肌肉标志物和纤维形成恢复,表明缺失的代谢物而非 PFKM 蛋白本身是主要瓶颈。 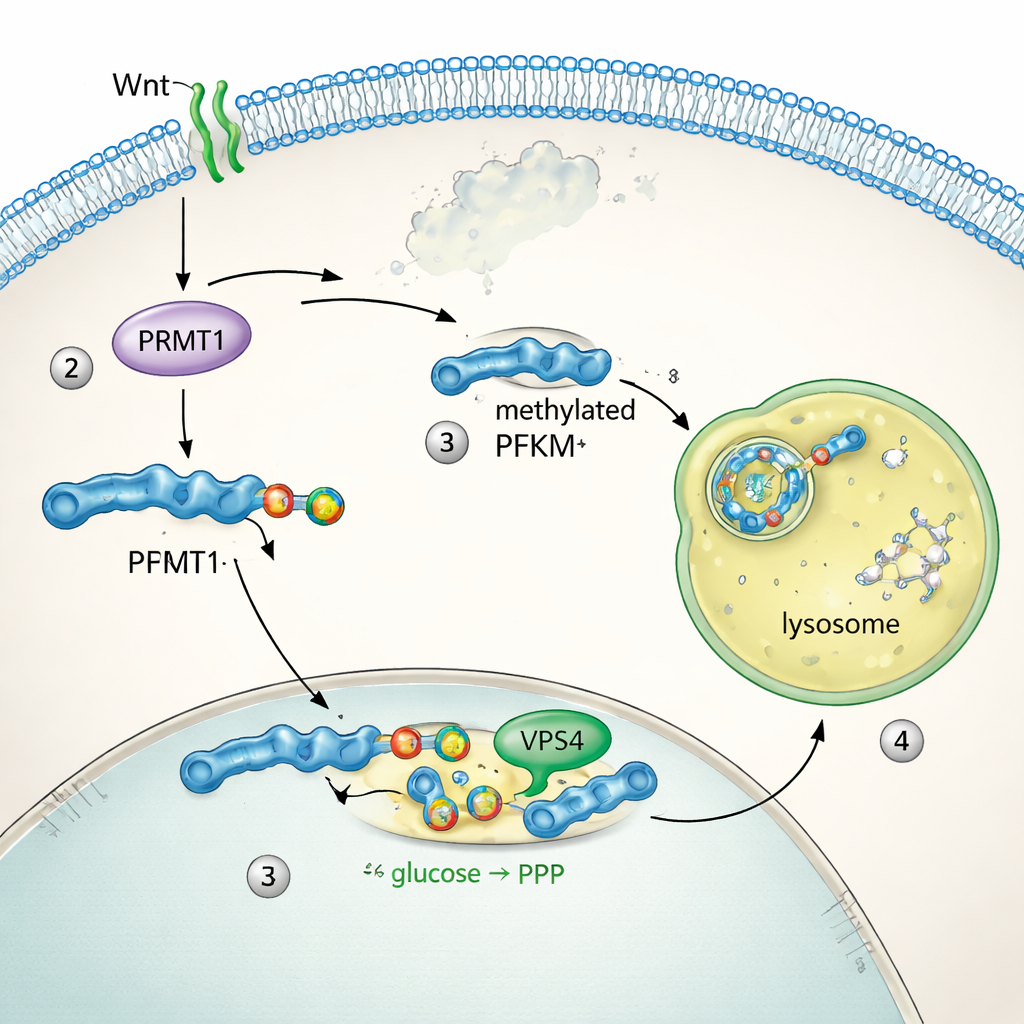
对肌肉健康与修复的启示
对非专业读者而言,核心信息是肌细胞并非简单地“多或少燃烧糖”;它们会根据生命阶段谨慎地在能量生产与细胞保护之间重新分配葡萄糖。PFKM 像一个可控阀门坐落在这一交汇处。在类干细胞中,Wnt 驱动的标记和溶酶体破坏 PFKM,将葡萄糖重定向到保护细胞并为未来生长做准备的途径。随着细胞成熟为能工作的纤维,它们重建 PFKM 储备并将代谢转向高输出能量利用。打破这种平衡(如罕见的 PFKM 缺陷病所示)会扰乱正常的肌肉发育。通过发现这一开关的分子调控点,该研究提示未来的疗法可能通过适度调整 PFKM 活性或补充合适的下游代谢物来微调肌肉再生,或在疾病和衰老中保护肌肉。
引用: Campos, M., Nguyen, S.T., Kong, X. et al. PFKM governs metabolic shifts throughout skeletal muscle differentiation. Nat Metab 8, 489–505 (2026). https://doi.org/10.1038/s42255-026-01457-4
关键词: 骨骼肌分化, 葡萄糖代谢, PFKM 酶, 戊糖磷酸途径, 溶酶体蛋白降解