Clear Sky Science · zh
来源于苔类植物的单体与二聚体光系统 I–LHCI 超复合体的结构研究
微小植物如何应对陆地生活
地钱类植物是最早从水域登陆的植物之一,远早于树木或开花植物的出现。为了在强光、干旱和紫外线等严峻环境中生存,它们必须重新设计细胞内那些微小的“太阳能电池板”。这项研究以近原子分辨率窥视了这些能量机构,揭示了原始陆生植物——地钱 Marchantia polymorpha——如何布置其光能捕获装置,以及这种结构如何可能帮助植物征服干旱陆地。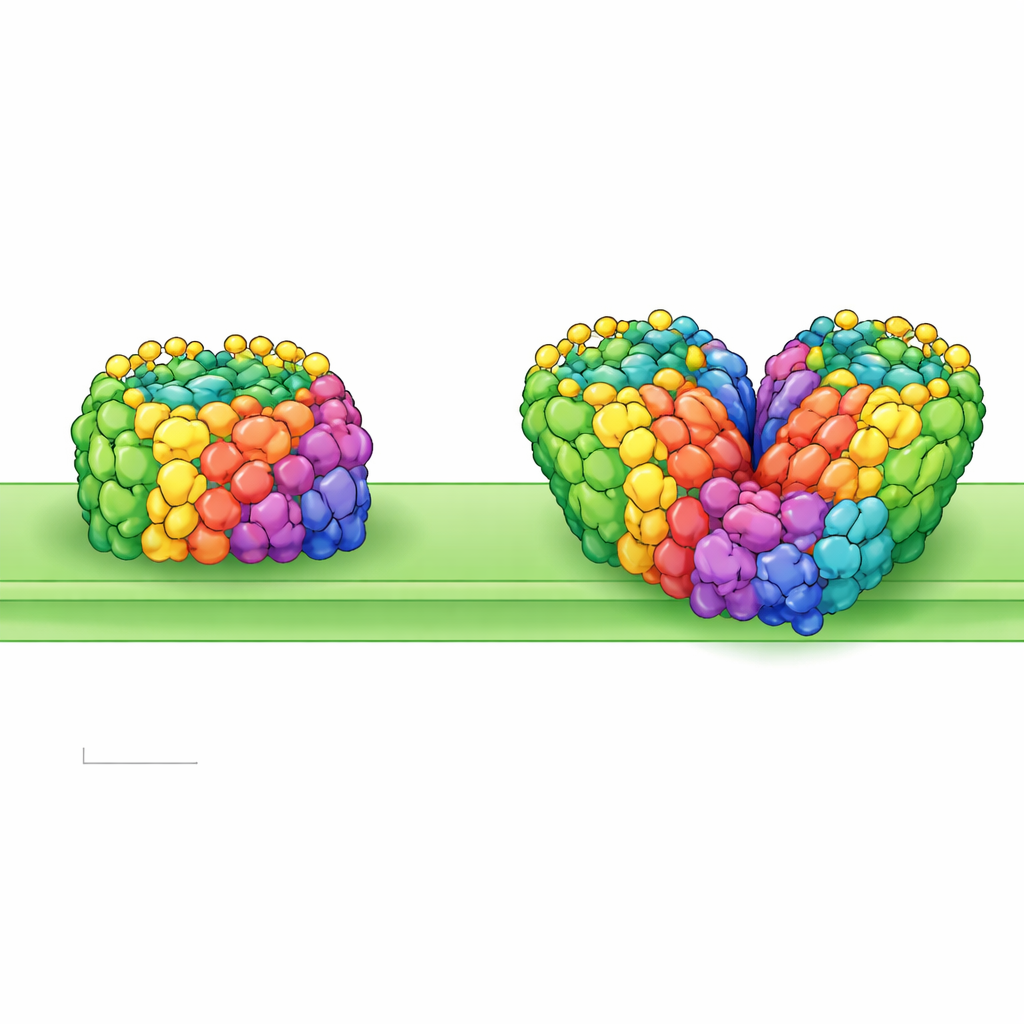
绿色细胞中的光能引擎
所有绿色植物都依赖称为光系统的微观机器将阳光转化为可用能量。其中最重要的之一是光系统 I,它通过一连串的色素和铁硫簇传递电子,最终为细胞的“化学电池”供能。环绕在该反应中心周围的是光捕获复合体,它们像天线一样捕捉多余的光并将能量汇入中心。二者共同形成嵌入叶绿体内膜的大型超复合体,即光系统 I–LHCI。尽管这一基本布局从细菌到开花植物都存在,但天线复合体的数量与排列随物种与环境而异,这暗示进化在应对昏暗林下、明亮田野或水中过滤光照等不同光环境时对该设计进行过反复改造。
以极高分辨率看到单体与二聚体
研究人员从 Marchantia 的类囊体膜中纯化出光系统 I–LHCI,并使用最先进的冷冻电子显微镜对颗粒成像,获得足以区分单个色素分子乃至许多水分子的分辨率。他们发现了两种主要形态:单一单元(单体)和两个单元结合在一起的结构(同源二聚体)。单体包含 13 个核心蛋白亚基和四个天线蛋白,每个天线均负载有叶绿素和类胡萝卜素以捕获光线。其整体形状与某些苔藓的相似,表明早期陆生植物间存在保守的构型蓝图。然而二聚体更为特殊:两个完整的单体面对面连接,但略有倾斜与扭曲,使整个组装看起来弯曲而非平坦。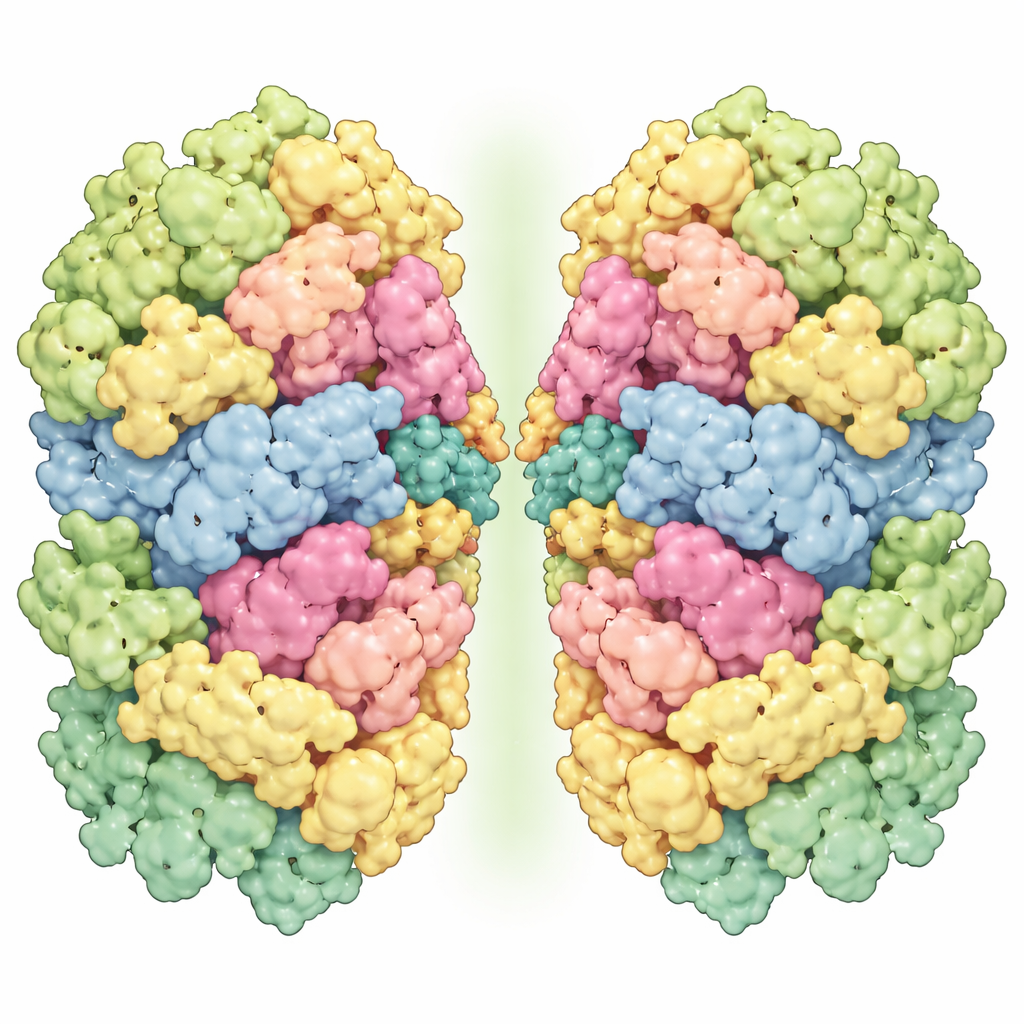
色素与能量流的精细布线
凭借这些清晰的电镜密度图,团队能够追踪几乎所有向反应中心传递能量的色素位置。他们确认了将电子从被称为 P700 的特殊叶绿素对运走的经典辅助因子链,并测量出与苔藓相比微小的位移——以埃为单位的分数。这些位移在实验不确定性范围内,但表明即使是密切相关的物种也会微调关键色素周围的局部环境。作者还记录了四个天线蛋白中各叶绿素分子的位置,以及这些蛋白环状区的细微变化如何产生或消失色素结合位点。通过理论计算,他们绘制出叶绿素之间可能的能量传递路径,并显示尽管二聚体中两个单体相互接触,但界面并不存在强烈的色素–色素耦合。换言之,二聚体似乎更像一种结构性排列,而非用于在搭档间共享光能的机制。
两个太阳单元如何相互黏合
地钱光系统复合体最显著的特征是其二聚体的连接方式。接触面涉及若干核心蛋白——PsaB、PsaM、PsaI、PsaG 和 PsaH——形成两处主要相互作用区。一处从膜的顶端延伸到底端,主要依赖 PsaM、PsaI 与 PsaB 之间紧密、排斥水分的接触,这表明 PsaM 对将两个单体锁定为一对至关重要。另一处位于类囊体基质侧,PsaH 跨越接触并触及相邻单体的 PsaG 及附近色素,帮助产生观察到的倾斜与扭曲。有趣的是,PsaM 在蓝藻、藻类与苔类中存在,但在被子植物中缺失;而 PsaG 与 PsaH 则在绿藻与陆生植物中保守。这一分布模式提示,地钱和苔藓保留了一种基于 PsaM 的较古老的光系统 I 二聚化方式,而这一方式在更高级的植物中要么丧失要么被改造。
这对植物进化意味着什么
通过提供来自陆生植物的首个高分辨率光系统 I–LHCI 二聚体结构,这项工作勾勒出光合“硬件”进化的关键一步。地钱似乎处在水生绿藻(可形成各种光系统 I 寡聚体)与维管植物(目前已明确为单体)之间的桥梁。Marchantia 中相对脆弱的二聚体由 PsaM 与 PsaH 稳定,支持了这样一个观点:在潮湿、低光的栖息地中,二聚体形式曾被青睐,但随着植物向更明亮、更干燥的陆地环境适应,这种形式变得不必要甚至不利。理解这类谦逊地钱中的结构解决方案,有助于阐明早期植物如何调整其太阳能装置以应对陆地生活的挑战。
引用: Tsai, PC., La Rocca, R., Motose, H. et al. Structural study of monomeric and dimeric photosystem I-LHCI supercomplexes from a bryophyte. Commun Biol 9, 146 (2026). https://doi.org/10.1038/s42003-026-09631-w
关键词: 光系统 I, 光能捕获, 地钱类植物, 冷冻电子显微镜, 植物进化