Clear Sky Science · zh
听觉皮层调节大鼠叫声时长
为什么大鼠叫声对理解言语很重要
当我们说话时,大脑会不断监听自身的声音并实时做出微小调整。这种自我监控对于清晰的言语至关重要,但在脑电路层面上它如何实现仍在探索中。在这项研究中,研究者转向大鼠及其超声叫声,提出了一个简单却根本的问题:负责听觉的脑区是否也参与控制叫声的持续时间?通过结合精确的脑电记录、局部药物注射和背景噪声,他们表明听觉皮层不仅是被动地聆听——它还主动塑造发声输出。
一边听一边发声
大鼠使用高频超声发声来传达情绪和社交信息。这些叫声依赖于横跨皮层、中脑和脑干的网络。为探究听觉皮层如何参与,研究团队电刺激了一个已知可触发叫声的中脑区域——导水管周围灰质。在麻醉状态下,当大鼠产生一系列超声声音时,研究者使用高密度 Neuropixels 探针记录了数百个听觉皮层神经元的活动。他们还将同样的叫声回放给动物听,从而可以直接比较自发发声时和被动听到时的大脑活动。
神经元响应叫声的五种方式
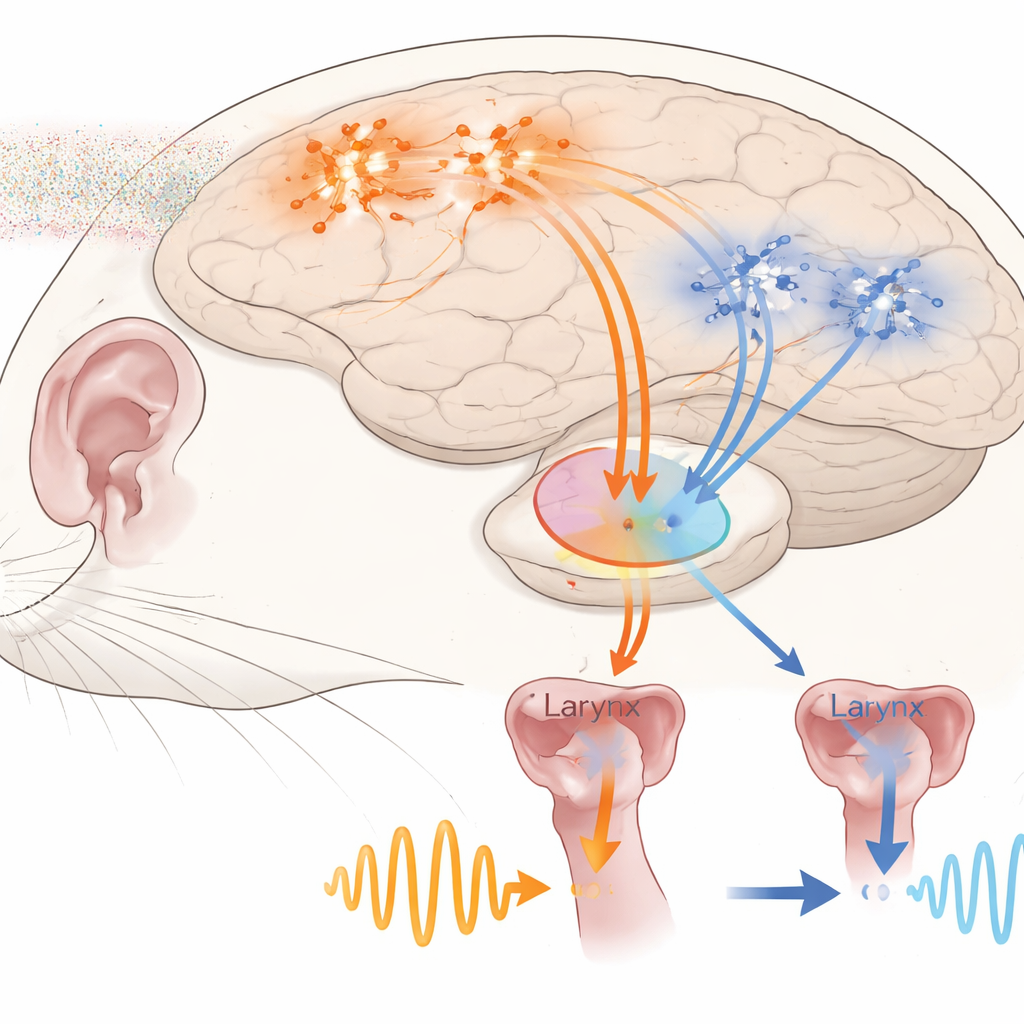
记录显示,听觉皮层的神经元并非都以相同方式工作。作者根据在叫声起始和结束时它们放电的变化,将细胞分为五种功能类型。有些细胞在叫声前激发(“预叫”细胞),有些在叫声开始时活动跃升(“起始激活”),也有些在起始时活动下降(“起始抑制”)。另外几组则表现出围绕叫声结束缓慢上升或下降的活动斜坡。重要的是,许多神经元在大鼠自己发声时对相同声音的反应与仅仅听到回放时不同。即便基线放电相似,自发发声的反应时序也更快,这表明来自运动相关的内部信号到达听觉皮层并在发声过程中对其进行特异性调节。
预测叫声时长的深层细胞
在这五类中,主要位于听觉皮层深层的起始抑制神经元尤为突出。作为一个群体,这些细胞在叫声开始前短短100毫秒窗口内的放电水平可以可靠地预测随后叫声会持续多久。更高或更低的预叫活动分别对应更长或更短的即将到来的叫声,这一关系跨动物成立,且并非由少数异常值驱动。同一群细胞还可以预测一个叫声序列会继续还是停止。仅利用叫声结束后短暂“预测间隔”内的活动,一个简单的机器学习分类器就能高准确率地判断是否会有另一次叫声跟随。这些发现表明,听觉皮层包含携带关于关键发声特征的预先信息的神经元,而不仅仅反映已经产生的声音。
转动皮层“旋钮”以延长或缩短叫声
为检验听觉皮层是否不仅提供信息而且具有因果作用,研究者直接改变了其活动。用促进抑制的药物肌松胆碱(muscimol)沉默听觉皮层会使总叫声时长变长并略微降低叫声音高,这一变化无法用生理盐水注射的对照来解释。相反,用阻断抑制输入从而激活局部网络的加巴嗪(gabazine)激活皮层则缩短了总叫声时长。在这种情况下,叫声音高大体保持不变。这些双向效应表明,更强的听觉皮层驱动倾向于抑制正在进行的发声,而降低皮层活动则让中脑的发声机制运行更久。团队接着探问,自然刺激——输送到耳朵的白噪声——是否会产生类似后果。
噪声作为对发声输出的自然刹车
当在嘈杂环境中触发叫声序列时,大鼠总体上发出更少且更短的叫声,音高更高且响度略有增加(与安静试验相比)。增大噪声强度在大多数动物中加强了这些变化,特别是总叫声时长的缩短和音高的上升。时机也很重要:噪声在叫声之前出现往往会延长该叫声,但与叫声重叠的噪声会缩短它并覆盖掉先前的延长效应。由于噪声会驱动整个听觉通路的活动,这些结果暗示声音诱发的听觉皮层及相关区域的激活会反馈到发声中心,并以梯度化、依赖强度的方式重塑叫声时序和声学特性。
这对言语与发声控制的意义
综上,这些实验表明大鼠的听觉皮层并非被动的麦克风,而是发声生成的主动参与者。特定的深层神经元携带关于叫声将持续多长时间以及是否会有后续叫声的预先信息,而听觉皮层总体活动的变化可以延长或缩短整体发声输出。背景噪声产生了类似的调整,暗示动物可能采用一种通用策略,通过调整叫声时长、音高与响度来应对嘈杂环境。这些发现拓宽了我们对感觉与运动系统如何相互作用的理解,并为研究支持灵活、基于反馈的发声行为——以及最终的人类言语——的脑电路提供了一个可行的模型。
引用: Tang, W., Concha-Miranda, M. & Brecht, M. Auditory cortex modulates call duration in rats. Commun Biol 9, 353 (2026). https://doi.org/10.1038/s42003-026-09608-9
关键词: 听觉皮层, 发声控制, 超声通话, 噪声诱导的发声变化, 大鼠交流