Clear Sky Science · zh
兴奋性GABA受体塑造秀丽隐杆线虫的运动回路组织
一只微小的线虫如何改写教科书规则
在生物课堂上,一种叫做GABA的大脑化学物质通常被介绍为制动器:它使神经元安静下来,帮助控制活动。对微小圆形线虫秀丽隐杆线虫(Caenorhabditis elegans)的这项研究颠覆了这一规则。作者展示了在这种简单动物中,GABA也可以像加速器一样作用于运动,通过推动特定的运动神经元来驱动精确的向后爬行。通过追踪哪些细胞产生哪些受体以及这些细胞如何相互连接,他们揭示了一种出人意料的巧妙方式:小型神经系统如何从有限的零件中挤出更多的灵活性。
简单神经系统中的多样通道阵容
秀丽隐杆线虫只有302个神经元,却能表现出令人惊讶的多种行为,从探索和逃避到协调进食和产卵。这种多功能性的一个重要来源是配体门控离子通道——在结合像GABA或乙酰胆碱等化学物质时开启的小型蛋白质孔道。与人类相比,这种线虫拥有数量过剩的此类通道:共计102个lgc基因。许多通道表现异常,对意外的化学物质有反应或允许正电荷而非负电荷流动。其间有一些罕见的GABA受体会兴奋而非沉默其所在细胞。直到现在,这些特殊受体在控制线虫身体弯曲的运动回路中部署在哪里仍不清楚。
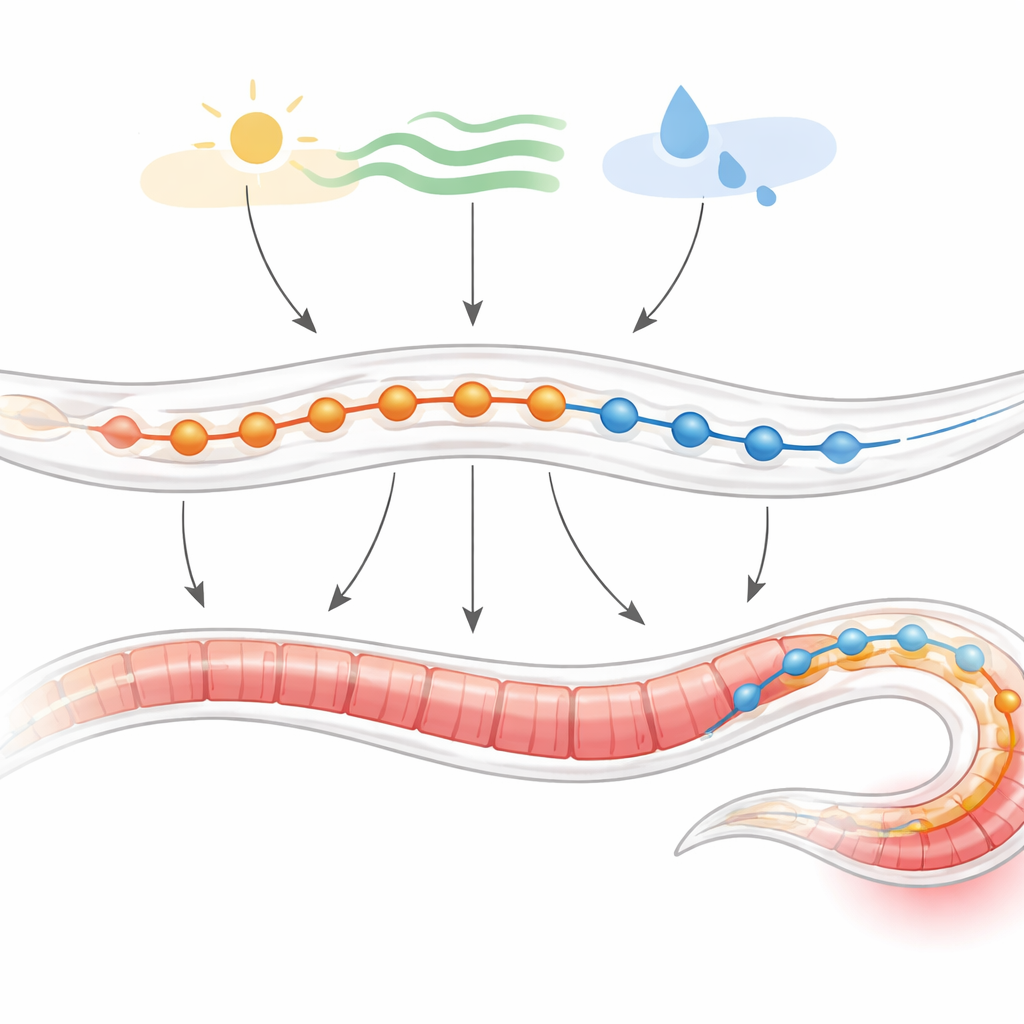
寻找运动控制的热点
研究人员挖掘了大型单细胞RNA测序图谱,这些图谱登记了线虫神经系统中各单个神经元中活跃的基因。他们发现,整个lgc家族在运动神经元中特别活跃,且在产生爬行节律性体波的那些神经元中活性最强。在这些与运动相关的运动神经元中,编码GABA受体的基因尤为突出。利用高分辨率的运动神经元亚型地图进一步观察,显示GABA受体分布在三个关键群体中:驱动向后运动的A型神经元、驱动向前运动的B型神经元,以及提供GABA信号的D型神经元。这些类别中超过一半的细胞携带至少一种GABA受体基因,表明GABA在塑造运动方面具有广泛且细致的作用。
兴奋性GABA在尾部的集中分布
并非所有GABA受体的行为都相同。线虫中的大多数是传统的抑制性受体,但有两个,称为EXP-1和LGC-35,允许正电荷流动,从而兴奋神经元。通过按每个运动神经元表达的GABA受体基因对其分类,团队发现许多A型和B型神经元在抑制性和兴奋性受体之间混合,使GABA有可能在不同情境下既抑制又增强活动。在驱动向后爬行的A型神经元中出现了一个显著的模式:越靠近尾部的神经元越可能携带兴奋性GABA受体。特别是LGC-35以及在最末端的细胞中出现的EXP-1,在这些后端神经元中富集,而两者常常避免出现在同一细胞中。这沿身体形成了一个兴奋性的空间梯度,使尾部对GABA特别敏感。
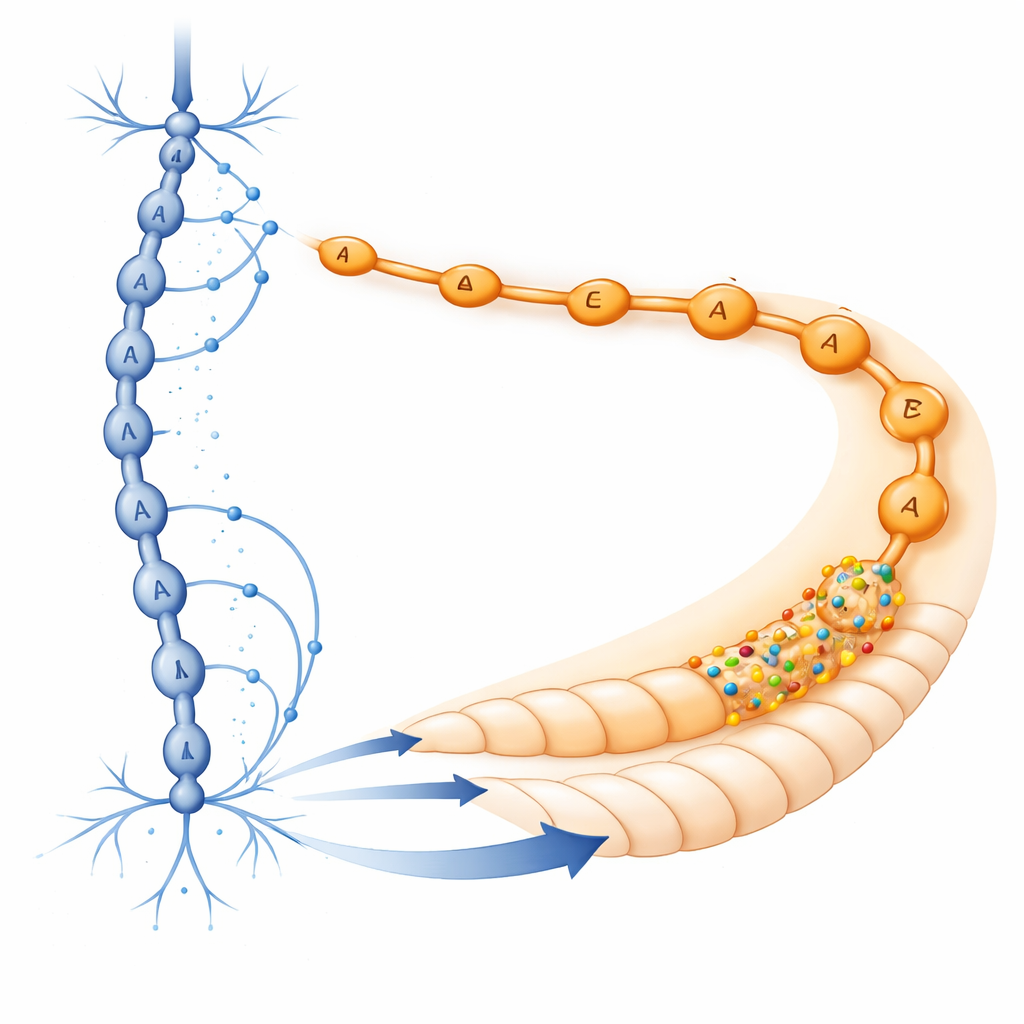
改写GABA经典图景
为理解这一分子模式如何与线虫的实际连线图相联系,作者转向基于电子显微镜的完整连通组。他们关注D型神经元——运动系统中主要的GABA释放细胞。这些神经元沿体节形成有序的突触链,连接到沿体的A型和B型运动神经元,其中背侧D型细胞主要与A型神经元相连。当将这一解剖图与受体表达数据叠加时,出现了清晰的图像:D型神经元向尾部区域的A型神经元发送GABA,而这些A型神经元富含兴奋性受体。早期工作还表明LGC-35还可以拾取从突触外溢出的GABA,进一步扩大其影响范围。综合来看,这些发现暗示了长期被认为纯粹抑制性的GABA系统实际上内置有一个在特定位置部署的兴奋性成分。
这对运动如何被引导意味着什么
对于非专业读者,关键结论是:这只微小线虫的运动方向并非由简单的开关控制,而是由沿体安置精心排列的化学“调节钮”模式控制。同一种信号分子GABA,依据每个细胞展示的受体类型以及其在头尾轴上的位置,可以使一些运动神经元减速而使另一些加速。通过将兴奋性GABA受体集中于驱动向后运动的尾部神经元,线虫似乎为尾部先行的运动(例如快速后退)提供了额外的推动力和精细控制。这项工作提出了一个更广泛的原则:即便是非常小的神经系统,也能通过以不同方式重复使用共同的化学物质、仅通过改变受体的摆放位置,来实现复杂且具方向性的行为。
引用: Wang, X., Mizuguchi, K. & Hashimoto, K. Excitatory GABA receptors shape locomotor circuit organization in C. elegans. Sci Rep 16, 9407 (2026). https://doi.org/10.1038/s41598-026-39358-x
关键词: 秀丽隐杆线虫运动, GABA受体, 运动回路, 单细胞转录组学, 神经连通组