Clear Sky Science · zh
原生β-羧体组装中生物分子冷凝体形成的阶段
为什么微小的碳工厂很重要
地球的食物链依赖光合作用,但负责从空气中固定二氧化碳(CO2)的关键酶 Rubisco 反应缓慢且易被氧气误识别。蓝藻——这些微小的光合微生物——早已通过构建称为羧体的蛋白“微工厂”解决了这个问题,这些结构在 Rubisco 周围富集 CO2。本研究解析了由 ApN 和 CM 蛋白组成的关键接头复合体如何分阶段自组装,以协助构建功能性的羧体。理解这一组装过程可为将类似的 CO2 提升系统引入作物植物并提高产量提供指导。
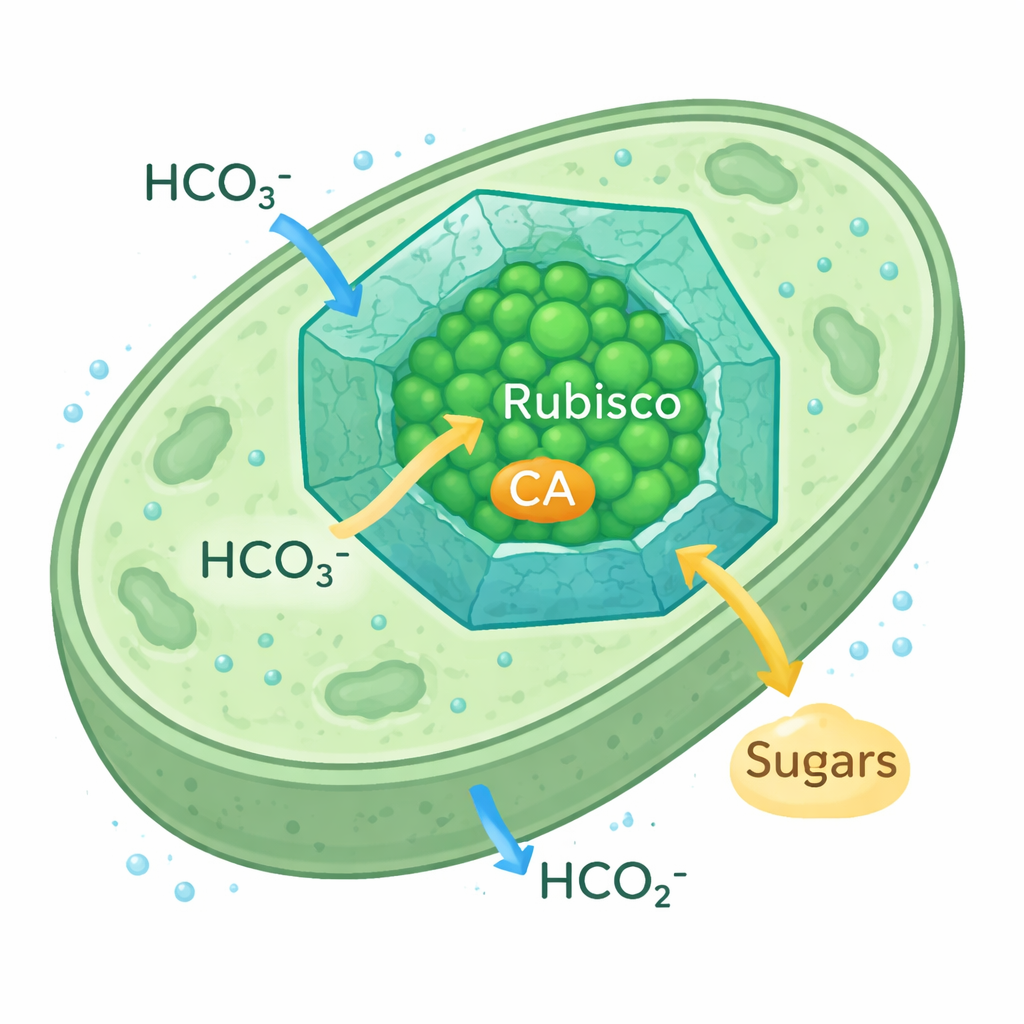
微生物如何增强碳捕获
蓝藻羧体是细胞内微小的二十面体蛋白隔室。它们的蛋白壳允许碳酸氢根离子进入,同时减缓 CO2 的逸散,在 Rubisco 及其辅助酶碳酸酐酶(CA)周围形成 CO2 富集区,碳酸酐酶将碳酸氢根转换为 CO2。这一 CO2 浓缩机制使蓝藻即便在环境 CO2 稀缺时也能繁荣生长。由于羧体中使用的 Rubisco 形式与植物 Rubisco 相似,研究者将其视为向植物叶绿体重新工程以更有效捕碳的现实模型。
与壳层相连的拼图块
在硬壳形成之前,蓝藻先构建一个柔软、类似液滴的“前羧体”冷凝体。早期研究显示,一个名为 CM 的支架蛋白将 Rubisco 和 CA 带入该液滴。缺失的一环是壳层接头蛋白 ApN 如何加入该结构并随后将其连接到壳蛋白上。作者发现,当单独表达 ApN 时,它形成一个四聚体,且不与其他羧体成分相互作用。在这种状态下,ApN 无法被 Rubisco、CM 和 CA 拉入液滴,提示在实际羧体组装过程中必须采用 ApN 的另一种构象或组合形式。
共同构建的接头与氧化还原开关
通过观察蓝藻的基因排列,团队注意到 apn 位于同一操纵子中紧随 cm 之后,因此这两种蛋白通常会一起合成。模拟这一点,他们在细菌中从同一段 DNA 共同表达 ApN 和 CM。在这些条件下,ApN 不再形成独立的四聚体。相反,三个 ApN 亚基与一个 CM 亚基组装成特定的四元接头复合体,记作 (ApN)3:CM。该异源复合体能够容易地与 Rubisco、CA 和 CM 结合并加入冷凝体。结构建模和冷冻电子显微镜显示,ApN 与 CM 通过精细契合的“舌槽”界面相互作用;当界面中的关键 ApN 残基被突变时,该接头无法形成,细胞也无法构建出正常的羧体。
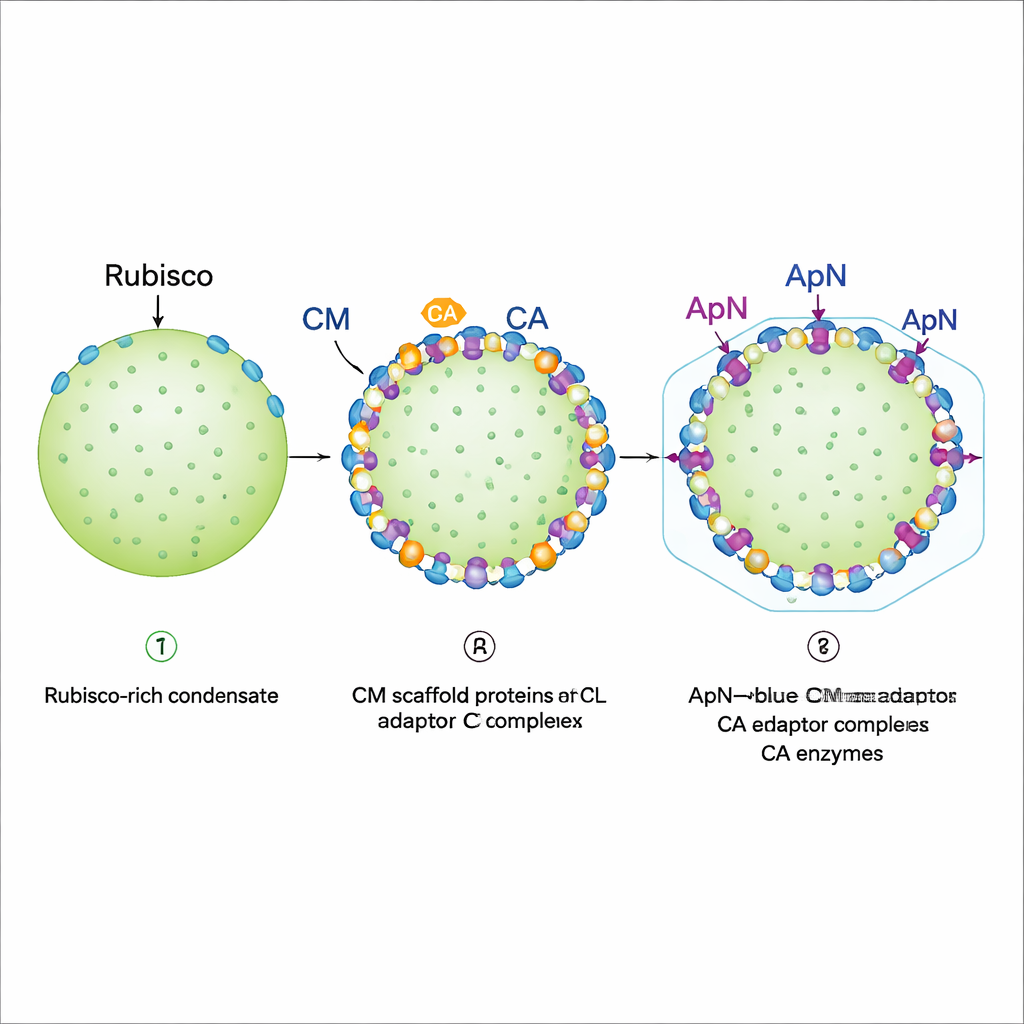
定位在边缘并为壳层做准备
对重构液滴的荧光成像揭示出分步的组装程序。首先,Rubisco、全长 CM 和一种短截的 CM 变体形成一个密集的冷凝体,表现出凝胶般的性质:Rubisco 基本被锁定在原位。随后加入 CA 和 (ApN)3:CM 复合体。只要 CA 先到或与 (ApN)3:CM 同时到达,这两者就倾向于沈积在现有液滴的外缘。这样的边缘定位至关重要,因为 ApN 的尾部带有一个“包封肽”,可结合壳蛋白。作者还发现一个内建的氧化还原开关:随着成熟中羧体内部环境变得更具氧化性,ApN 中两个保守的半胱氨酸残基可被氧化,促使接头从 (ApN)3:CM 四聚体向 (ApN)2:CM 三聚体重构。在活体蓝藻细胞中,改变这些半胱氨酸会减缓生长并产生体积过大的但仍能工作的羧体,突显了它们在微调成熟过程中的作用。
为改良作物提供的启示
对非专业读者来说,这些分子细节看似晦涩,但其要点很简单:时序与协同很重要。研究表明,必须与 CM 共同生成 ApN,才能避免其形成无用的自聚体,而是成为能够停驻在前羧体表面、准备招募壳层的接头。随后一个对氧化还原敏感的步骤随着隔室闭合及内部化学变化帮助重塑该接头。对于希望将蓝藻 CO2 浓缩机制移植到作物中的植物工程师而言,这项工作强调仅转移单个部件并不足够;还必须尊重在自然中引导羧体组装的顺序、比例和化学触发条件。
引用: Zang, K., Hong, X., Nguyen, N.D. et al. Stages of biomolecular condensate formation in pro-β-carboxysome assembly. Nat. Plants 12, 447–464 (2026). https://doi.org/10.1038/s41477-026-02227-6
关键词: 羧体组装, 蓝藻二氧化碳浓缩机制, Rubisco 冷凝体, 生物分子冷凝体, 合成光合作用