Clear Sky Science · zh
烟曲霉的光解酶/隐花色素感知氧化应激并在细胞核与线粒体间穿梭
真菌如何感知光与应激
阳光维持植物和真菌的生命,但同时也带来危险,例如造成 DNA 损伤和产生有害的活性氧化物。本研究探讨了常见霉菌烟曲霉(Aspergillus nidulans)如何利用单一蛋白 CryA 同时感知光和氧化应激并保护自身。理解这一“二合一传感器”不仅加深了我们对微生物如何应对恶劣环境的认识,也可能有助于阐明细胞如何在细胞核与能量产生的线粒体之间协调信号。
一个蓝光修复酶的隐藏角色
CryA 属于一类以修复紫外线引起的 DNA 损伤而著称的蛋白。这些蛋白称为光解酶与隐花色素,利用吸光分子(类黄酮)吸收蓝光并修复断裂的 DNA 碱基。研究者确认 CryA 具有典型的 DNA 修复酶结构,能够结合常见的光捕获辅因子,并在进化树上与已知光解酶分为一簇。单凭这些,CryA 看起来像是常规的修复工具,然而早期研究提示它也影响真菌的发育,表现更像受光调控的开关而非单纯的分子修复器。
光响应基因的主控调节器
为揭示 CryA 的调控作用,研究团队追踪了其在细胞中的定位以及改变其含量对真菌生长的影响。他们发现在正常条件下 CryA 积累在储存 DNA 的细胞核中。删除 cryA 基因时,真菌产生更多的有性结构;而强制过表达 CryA 几乎完全抑制了常见的无性孢子的形成,留下苍白、绒毛状的菌落。基因表达检测显示,许多光激活和发育相关基因在缺失 CryA 时被过度激活,而在 CryA 过量时则被弱化。综合这些结果表明 CryA 是一个负反馈元件:光增加 cryA 水平,CryA 随后进入细胞核并抑制光和发育诱导的基因,防止反应失控。 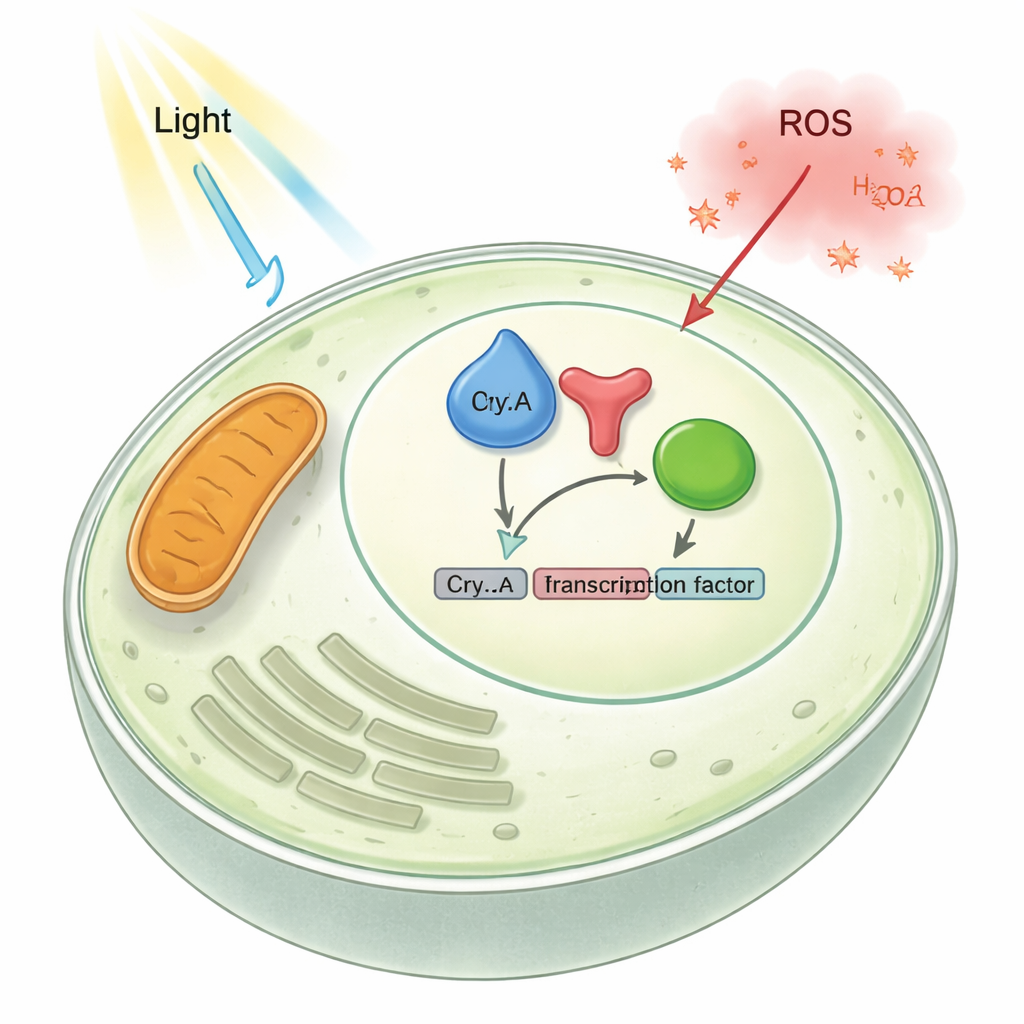
与主要光与应激通路的串扰
该真菌还依赖另一种光感受器——红光感受器光敏素(FphA),以及以转录因子 AtfA 结束的应激通路。通过活细胞与纯化蛋白的蛋白—蛋白相互作用测试,研究者显示 CryA 在细胞核中与 FphA 和 AtfA 都发生物理结合。当删除 cryA 时,通过光敏素介导的红光激活基因表达更强;而过表达 cryA 时,这些基因更难被激活。染色质实验表明,在缺失 CryA 的情况下,一个关键的光响应基因带有更多的激活性组蛋白标记,提示 CryA 通常抑制由光敏素驱动的染色质开放活性。实际上,CryA 既遏制光感受器也抑制下游的转录因子,作为光与应激信号的共同刹车。
快速的应激传感器并跃迁至线粒体
氧化应激——例如过量的过氧化氢等活性氧——对细胞构成持续威胁。作者发现,这类应激与光一样会提升 cryA 的表达。显著的是,当加入过氧化氢时,CryA 在不到一分钟内从细胞核移动到线粒体。这一跃迁依赖于蛋白 N 端的一段短而柔性的延伸片段,尤其是其中的一个半胱氨酸残基。当该半胱氨酸被替换为其它氨基酸时,CryA 在应激下无法离开细胞核。截短 N 端尾部会使 CryA 永久定位于线粒体。这些工程化菌株对氧化剂的反应不同:仅位于细胞核或仅位于线粒体的 CryA 会改变真菌对过氧化氢和对苯二酚(menadione)的耐受性,并重塑抗氧化基因的表达。研究结果表明 CryA 不仅是感受应激的传感器——它还可能帮助协调线粒体与细胞核之间的通信,使抗氧化防御与损伤类型和程度相匹配。 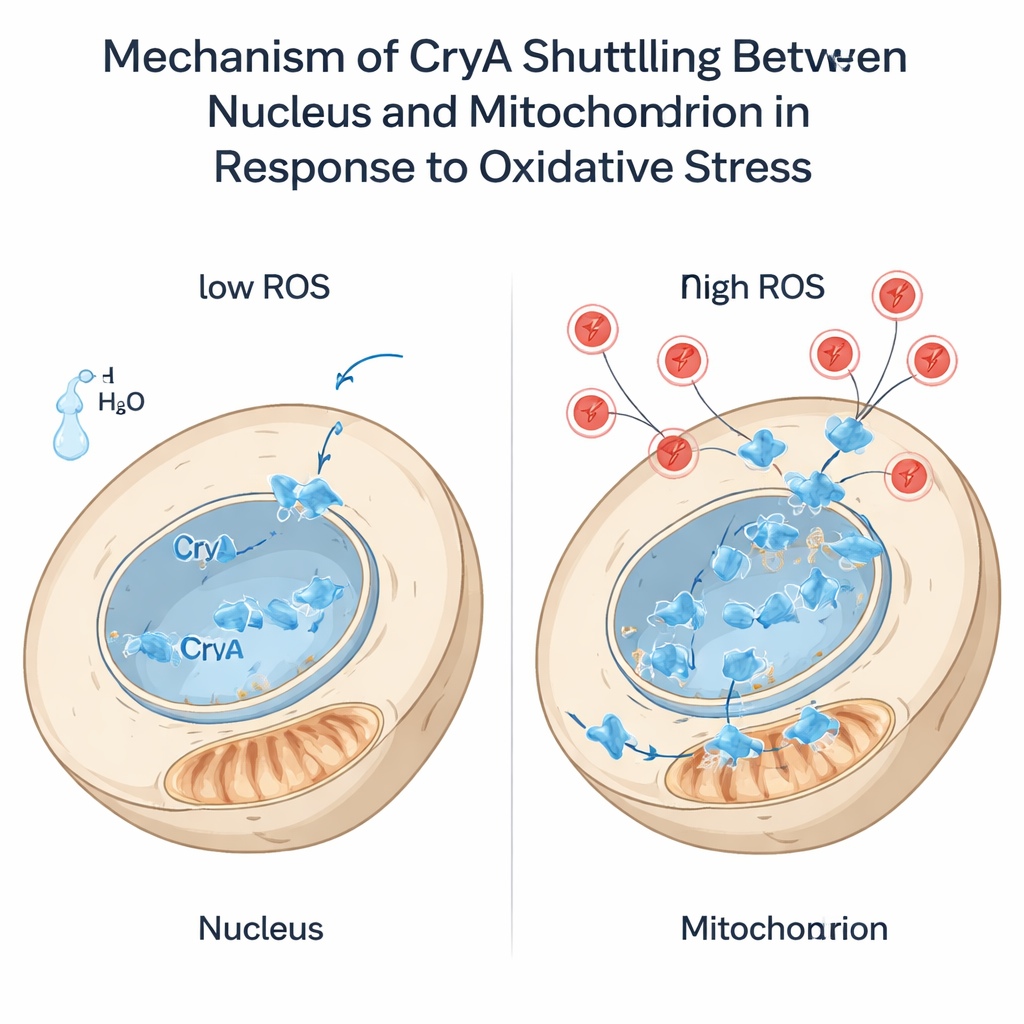
这项研究的重要性
对非专业读者来说,CryA 可被视为细胞交通警察,既监视外界的光照,又感知内部的氧化应激,然后决定何时放慢生长、发育和基因活性。通过在细胞核与线粒体间穿梭并接入主要信号通路,CryA 防止真菌对光或应激反应过度,同时仍能允许快速的保护性反应。许多生物体中存在类似的蛋白与机制,因此这项工作为理解活细胞如何将环境线索与内部损伤信号整合以在变化的世界中生存提供了重要视窗。
引用: Landmark, A., Rudolf, T., Hundshammer, K. et al. The photolyase/cryptochrome of Aspergillus nidulans senses oxidative stress and shuttles from nuclei to mitochondria. Nat Commun 17, 1483 (2026). https://doi.org/10.1038/s41467-026-69403-2
关键词: 光感应, 氧化应激, 隐花色素, 线粒体, 真菌发育