Clear Sky Science · zh
AACRNL 从毒力因子进化为在自由生活真核生物中推动基因组扩张的表观遗传寄生体
当古老毒素学会新把戏
我们的基因组并非寂静的说明书。它们是拥挤的竞技场,基因、病毒和可移动的 DNA 不断争夺空间与控制权。这项研究揭示了这一搏斗中的一个令人意外的参与者:一种起初作为微生物攻击宿主的毒素而存在的蛋白质,但后来在自由生活的动物体内被重新利用,成为一种帮助自身在基因组中扩张足迹的“表观遗传寄生体”。
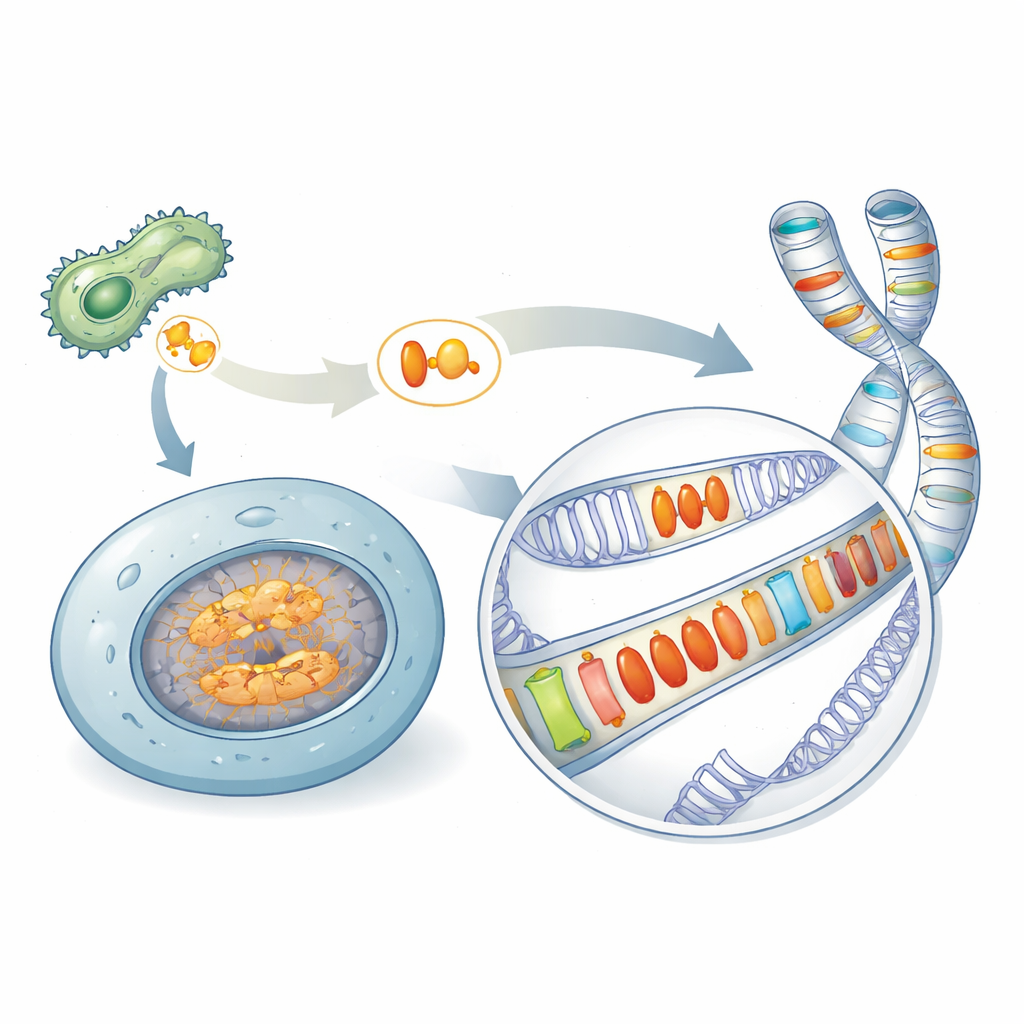
微生物武器的隐秘遗产
许多致病微生物依赖特殊蛋白质(称为效应子)来破坏其感染对象的防御系统。一个著名的家族,称为 Crinklers,曾被认为只存在于病原体中。通过扫描多种物种,作者们在意想不到的生物中发现了相关蛋白——在此归为 AACRNL 名下——包括海绵、珊瑚、海胆和一些非典型病原体的硬骨鱼类。这些 AACRNL 蛋白保留了类似毒素的核心,能够化学修饰其他蛋白质,但其前端用于病原体侵入宿主细胞的“传递”部分已大体崩解。这种结构变化暗示其功能已从攻击其他生物转向在宿主自身细胞内发挥作用。
基因组中的自私开关
在硬骨鱼中,AACRNL 基因以多个拷贝分布在不同染色体上。通过比对邻近的 DNA 片段,研究者表明一些 AACRNL 衍系已停止自我复制,而另一些则借助邻近的跳跃 DNA 片段——转座子——大量扩增。转座子像基因组穿梭器:当它们移动或复制时,能把 AACRNL 一并带走。其结果是在转座子序列中镶嵌出一组重复的 AACRNL 拷贝,这种拼凑式的分布是基因为自身利益而扩散,而非为宿主服务的明显特征。
打破基因组的静音按钮
通常,细胞会严密束缚这类可移动 DNA。其中一项关键防护是存在于 DNA 包装蛋白上的化学标记 H3K27me3,这有助于将危险或噪音较大的区域保持关闭。作者发现,硬骨鱼中一种活跃的 AACRNL 形式,称为 AACRNLβ,能够化学修饰负责添加该抑制标记的酶 EZH2。当 AACRNLβ 改变 EZH2 时,其自身基因位点及邻近转座子的 H3K27me3 水平下降,局部 DNA 变得开放,AACRNLβ 及其转座子邻居的表达显著增强。实际上,该蛋白按下了自己的“开启”按钮,并解除对那些能将其带到基因组新位置的移动元件的刹车。
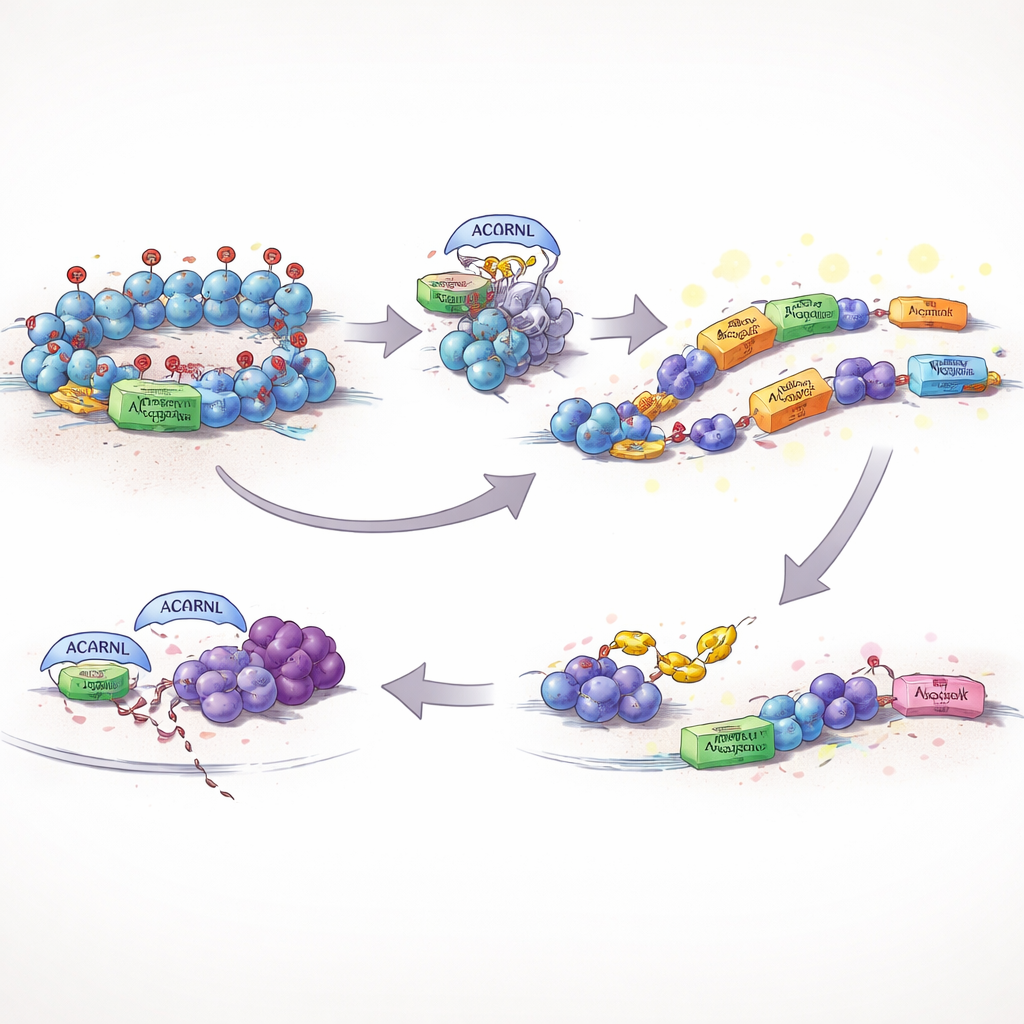
躲避免疫看守
释放转座子和异常的基因活动会冒引起免疫系统注意的风险。研究显示 AACRNLβ 也能应对这一问题。它向 TRAF6(一个在启动抗病毒和炎症信号中起中心作用的枢纽蛋白)添加了一个小的化学标记。被这种方式标记的 TRAF6 稳定性降低,更容易被细胞的废物处理机制标记并降解。随着这一信号枢纽被削弱,关键防御通路的激活减弱,形成一个更宽容的环境,使 AACRNLβ 及其相关转座子得以在被察觉的风险较小的情况下持续存在并复制自身。
细胞内部的脆弱军备竞赛
故事并未以 AACRNLβ 不受限制地扩张而结束。被 AACRNLβ 针对的同一个 TRAF6 蛋白反过来也能向 AACRNLβ 添加自身的标记,将这一类似寄生体的蛋白标记为待降解目标。这种你来我往类似分子层面的军备竞赛:AACRNLβ 利用源自毒素的化学活性去弯曲染色质状态和免疫反应以利于自身扩张,而宿主蛋白则通过降解它、使其活动变得零星且代价高昂来反击。作者认为正是这种拉锯允许前微生物武器以自私驻留者的身份在自由生活的基因组中存活,增加 DNA 含量并在内部重塑遗传冲突的规则。
这对我们理解基因组的重要性
对于非专业读者,关键讯息是:基因组不仅仅是为宿主生存优化的蓝图。它们也藏有机会主义者。这项工作表明,即便是经典毒素也可能被驯化为此类元件,利用其原有的破坏性化学特性绕过表观遗传锁和免疫警报,并在进化时间尺度内在基因组中扩散。理解这种隐秘冲突有助于解释基因组为何如此庞大、复杂且富于变化——并提示其他“退休”毒素可能在许多物种(或许包括我们自己)中悄然改写 DNA。
引用: Xu, T., Geng, S., Lv, X. et al. AACRNL evolved from virulence factor to epigenetic parasite driving genome expansion in free-living eukaryotes. Nat Commun 17, 2130 (2026). https://doi.org/10.1038/s41467-026-69012-z
关键词: 自私的遗传元件, 基因组进化, 表观遗传调控, 转座子, 先天免疫