Clear Sky Science · zh
乳酰胺酶进化中活性—稳定性权衡的动态特征
这对抗生素耐药为何重要
抗生素耐药常被视为一个黑盒:细菌“以某种方式”发生变化,药物失效。本研究为世界上最著名的耐药酶之一——TEM‑1 β‑内酰胺酶——打开了这只黑盒。该酶帮助细菌破坏类青霉素抗生素。通过观察该酶在进化以对抗更新型药物时其构象与运动如何改变,作者揭示了为何那些提高活性的突变常常削弱稳定性,以及进化如何找到巧妙的方式在两者之间取得平衡。
从青霉素克星到更广谱的药物破坏者
TEM‑1 原本擅长裂解较早的青霉素类抗生素,但对诸如头孢噻肟这类更大、更笨重的药物能力较弱。在许多临床菌株中,一个靠近酶活性位点的关键单点突变 G238S 经常出现。该突变显著提升了酶裂解头孢噻肟的能力,同时对其原先裂解青霉素的活性仅造成温和影响。作者表明 G238S 并非简单地扩大活性位点口袋;它重组了若干周围环和螺旋的运动方式,产生了一种新的功能性构象,更能容纳体积较大的药物。
将蛋白运动当作进化的调节旋钮
研究者使用先进的核磁共振(NMR)技术,测量了 TEM‑1 不同部位在从太秒(10⁻¹² s)到毫秒(10⁻³ s)量级的运动。野生型 TEM‑1 相当刚性,这有助于它高效处理原始底物。G238S 保留了大部分这种快速刚性,但在活性位点的多面“墙”中引入了更慢、经过精细调节的运动。这些运动足够快以配合酶促化学反应,却不会剧烈到破坏关键催化残基。结果是一个“优化的灵活窗口”:恰到好处的移动既能为头孢噻肟打开口袋,又能保持核心化学机制的正确对齐。
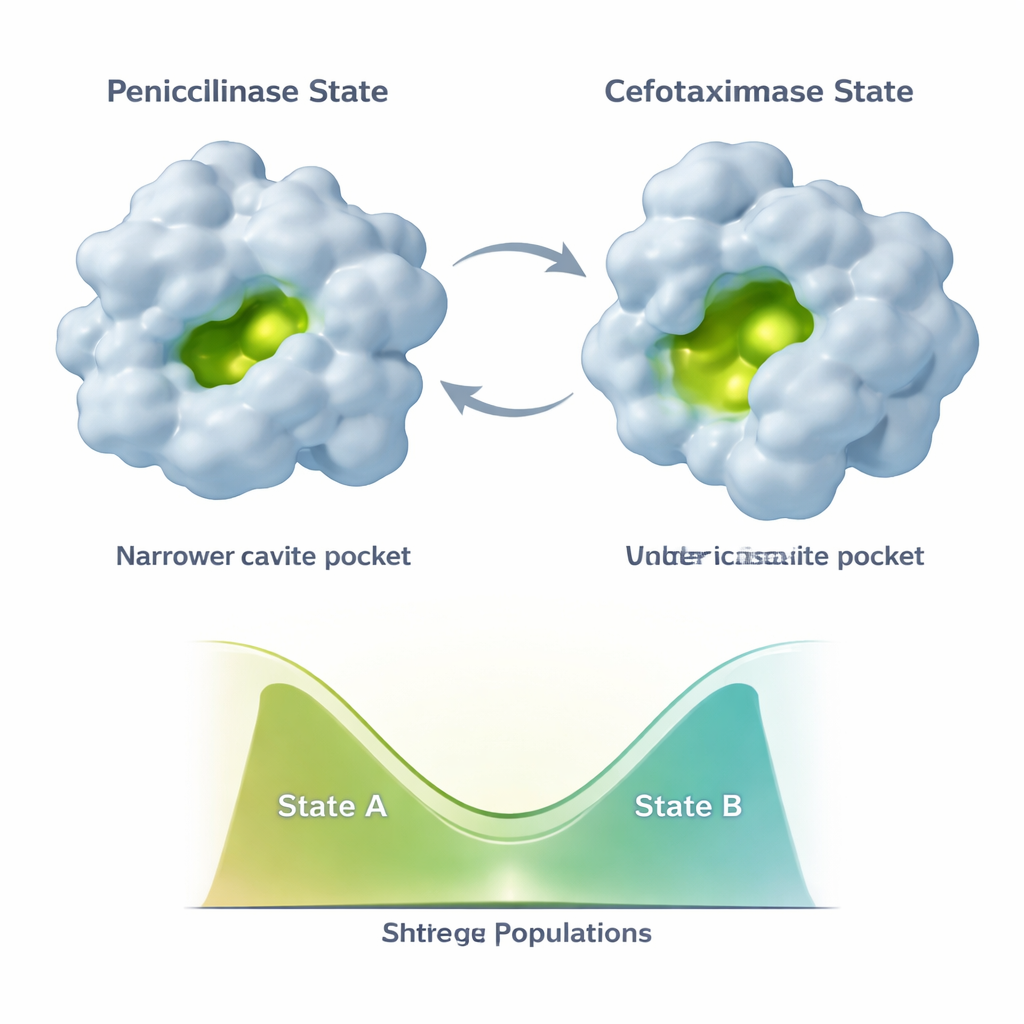
不是选择一种形态,而是在两种形态之间平衡
进化并没有把 TEM‑1 固定为单一新构象。相反,该酶至少采样两种主要构象:“青霉素酶”状态类似原始结构,而更开放的“头孢噻肟酶”状态更适合新药物。随后出现的额外突变,如 E104K 和 A42G,作用微妙。它们并非单纯使新的、利于头孢噻肟的构象更稳定,而是重新平衡这两种状态的混合比例。NMR 数据表明,活性位点的不同部分与支撑骨架可以沿着这两态连续体独立改变其占比。这导致了一组组合性的酶变体,每种变体在类青霉素与类头孢噻肟构象的混合比例不同,因此具有不同的催化特性。
隐藏的薄弱点与远端修补
提高活性的突变通常带来隐藏的代价:使蛋白质变得不那么稳定。研究团队没有只看完全展开,而是使用氢-氘交换结合质谱在短片段水平上映射局部稳定性。结果发现 G238S 不仅使附近的环不稳定化,还影响形成结构骨架的远端螺旋和β片层区。这些区域中有些与一个“隐秘”的变构口袋重合——蛋白核内不常被访问但能结合小分子并抑制活性的开口。G238S 使该口袋更容易打开,实际在酶中构建了一个轻度的自我抑制特征。随后出现的突变,尤其是 A42G,加固了这条被削弱的骨架网络,提高了三股相互作用螺旋周围的局部稳定性,而不消除有益的活性位点动力学。换句话说,进化修补的是远端的结构薄弱点,而不是撤销最初的创新。
这项研究揭示的进化策略
对非专业读者来说,核心信息是:像 TEM‑1 这样的蛋白并不是通过翻转一个简单的开关来进化出耐药性。每一次突变都在微调酶如何“呼吸”、如何弯曲,以及在不同工作构象之间如何分配时间。G238S 为酶打开了通向新功能的大门——裂解更新的抗生素——但同时也造成了局部脆弱性和部分自我抑制状态。继发突变像精心加固的修补,稳定骨架并微调旧构象与新构象之间的平衡,使酶既能保持活性又更为耐久。这种将运动与局部弱点视为与静态结构同等重要的动态进化观,可为设计未来更难被细菌“击败”的抗生素和靶向酶的药物提供指导。
引用: Arcia, E., Keramisanou, D., Jacobs, L.M.C. et al. Dynamic signature of activity-stability tradeoff in lactamase evolution. Nat Commun 17, 1884 (2026). https://doi.org/10.1038/s41467-026-68620-z
关键词: 抗生素耐药性, β-内酰胺酶, 蛋白质进化, 酶动力学, 蛋白质稳定性