Clear Sky Science · zh
DNA 甲基化与长非编码 RNA 在特定印记基因区控制异步 DNA 复制
为何细胞的 DNA 复制时间表很重要
每次细胞分裂时都必须复制完整的 DNA,但并非所有区域在同一时刻被复制。有些片段早期复制,另一些则在晚期复制,遵循基因组的精确“时间表”。这篇论文探讨了为何在少数记忆父母来源的特殊基因簇中,来自母方和父方的拷贝在复制时间上不同步。理解这种异常的复制时序有助于阐明 DNA 上的化学标记和长非编码 RNA(lncRNA)如何组织基因组,并可能影响发育与疾病。
写在 DNA 上的亲本记忆
在哺乳动物中,一小部分“印记”基因域会根据来自母亲或父亲的不同而表现出差异。这种亲源效应由 DNA 甲基化控制——即添加到被称为差异甲基化区(DMR)的特定区域的微小化学标记——以及染色体折叠的变化。作者研究了小鼠胚胎干细胞,这些细胞类似于非常早期的胚胎细胞。他们比较了仅含母源基因组、仅含父源基因组和携带一套来自各方的正常杂交细胞。通过追踪细胞周期中 DNA 各片段的复制时间,他们绘制了全基因组的复制时序并聚焦于已知的印记区。
两个印记区打破常规
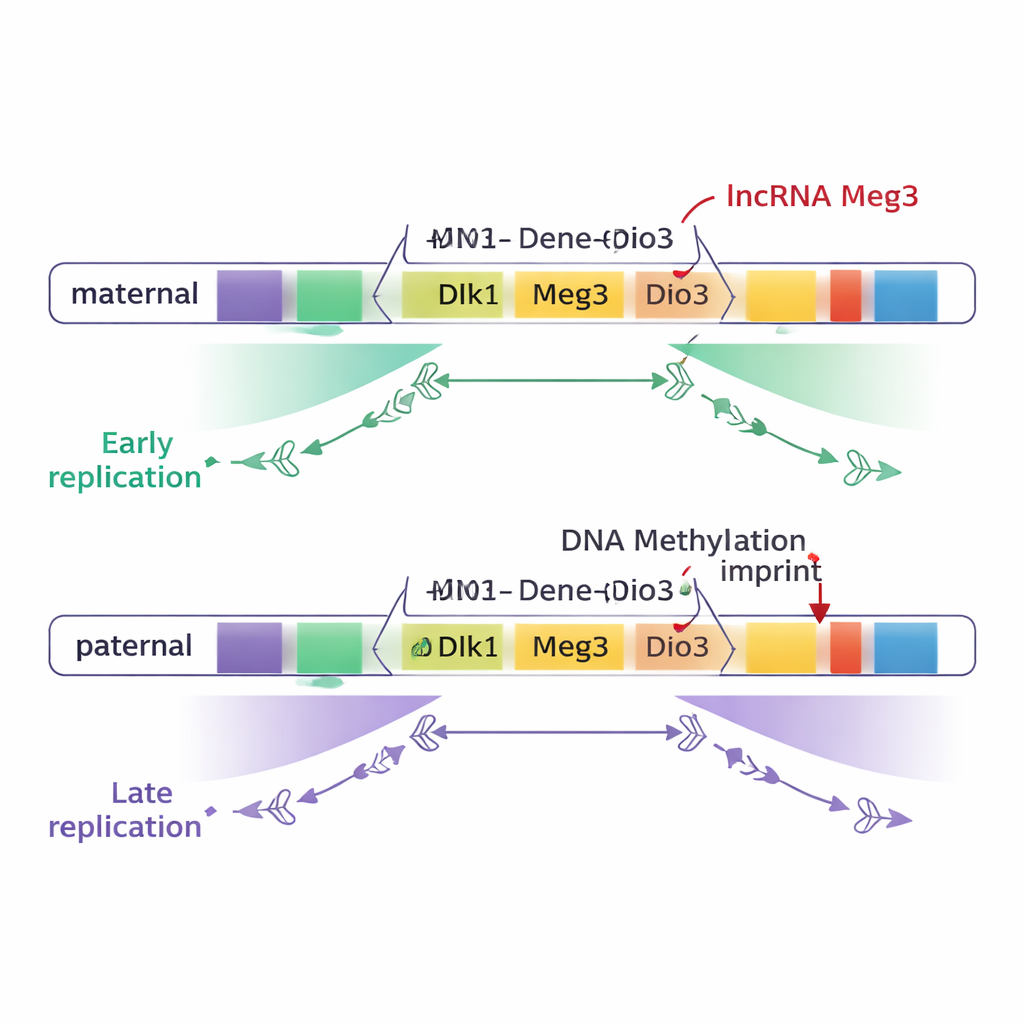
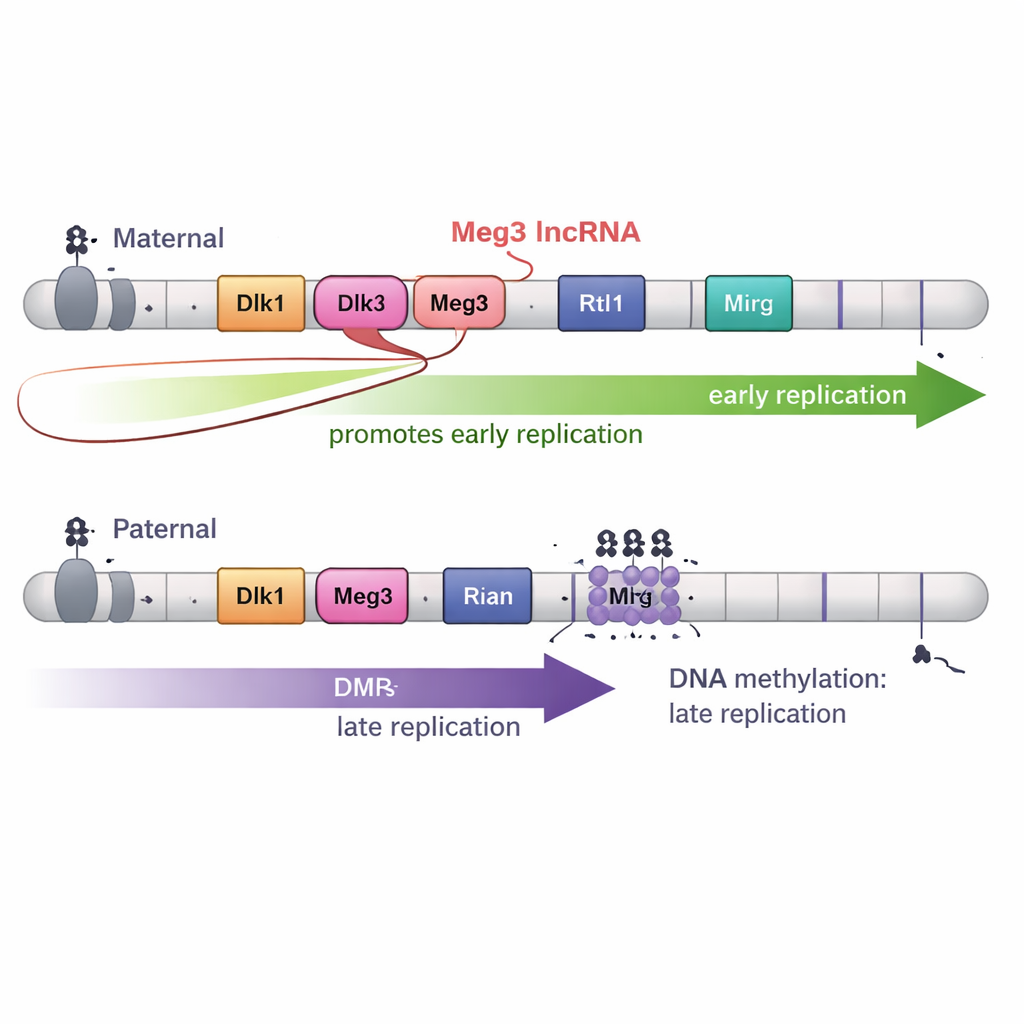
大多数基因组区域,包括大多数印记域,母源和父源染色体上的复制时间是一致的。然而,有两个大的印记区例外:染色体12上的 Dlk1–Dio3 域和染色体7上的 Snrpn 区。在这些区域,一大片 DNA——在 Dlk1–Dio3 约有 75 万碱基对——在一条亲本染色体上早期复制,而在另一条上晚期复制。关键的是,这种差异遵循亲本来源,而非品系背景:例如 Dlk1 和长非编码 RNA 基因 Meg3 的母源拷贝持续早期复制,而父源拷贝则晚期复制。
DNA 甲基化设定复制时序开关
为测试导致这一时序差异的原因,研究者工程化了胚胎干细胞,使 DMR 处的正常甲基化模式要么被抹去,要么被强制施加到两条亲本染色体上。当 Dlk1–Dio3 DMR 的两条拷贝都被甲基化时,该区域在两条染色体上都呈晚期复制;当两条拷贝的甲基化被大量去除时,同一区域在两条染色体上都呈早期复制。在 Snrpn 处进行的类似实验也导致复制时间差异的丧失。这些结果表明,亲本特异的 DNA 甲基化对于在这些域中产生母源与父源之间的早晚复制对比是必需的。
一条长 RNA 微调早期复制
然而,DNA 甲基化并非全部故事。Dlk1–Dio3 区还产生一条名为 Meg3 的长非编码 RNA,作为一个大型 RNA “多顺反子” 的一部分。在母源染色体上,未甲基化的 DMR 允许 Meg3 表达;在父源染色体上,甲基化使其沉默。通过创建精确的缺失以关闭 Meg3 而保持 DNA 甲基化不变,作者表明 Meg3 RNA 的丧失会使母源域的特定部分从早期向更晚的复制转移。换言之,Meg3 RNA 有助于促进母源染色体附近片段的早期复制,为 DNA 甲基化之上增加了第二层控制。
三维折叠与发育带来更多变数
鉴于基因组折叠常与复制时序相关,团队还高分辨率绘制了染色体的三维结构。出人意料的是,在 Dlk1–Dio3 区域,三维结构域的边界并不匹配早晚复制的界限。即使在甲基化模式被改变、复制时序随之改变时,基本的折叠单元——拓扑关联域(TADs)——也以不同方式移动。最后,当干细胞分化为神经祖细胞时,母源与父源拷贝之间显著的时序差异大部分消失,尽管印记标记和三维结构的许多方面仍然存在。这表明发育线索可以覆盖干细胞中观察到的那种特殊早晚复制模式。
通俗的结论
这项研究揭示,在少数记住亲本来源的特殊基因邻域中,细胞的 DNA 复制时间由 DNA 上的化学标记和长非编码 RNA 的组合所控制。印记控制区的 DNA 甲基化为每条亲本染色体设定了基本的早或晚复制时序,而 Meg3 lncRNA 进一步促使邻近的母源 DNA 更早被复制。这些效应在很大程度上独立于染色体的三维折叠。随着细胞在发育过程中分化,这种不寻常的时序会消退,表明它是多能干细胞状态的特征。总体而言,该研究解释了亲本的表观遗传“记忆”和非编码 RNA 如何在局部覆盖基因组通常的复制时间表。
引用: Imaizumi, Y., Charon, F., Surcis, C. et al. DNA methylation and lncRNA control asynchronous DNA replication at specific imprinted gene domains. Nat Commun 17, 1844 (2026). https://doi.org/10.1038/s41467-026-68558-2
关键词: 基因组印记, DNA 甲基化, 复制时间, 长非编码 RNA, 胚胎干细胞