Clear Sky Science · zh
独立shelterin成分对端粒酶招募与端点保护的控制
细胞如何保护染色体末端
每次细胞分裂时,染色体的末端——称为端粒——都会被逐渐缩短。为了避免丢失重要的遗传信息,细胞使用一种特殊酶——端粒酶,以及一组保护性蛋白来维持这些末端。本研究探讨了其中两种蛋白TPP1和POT1如何在允许端粒酶进入以重建端粒与将染色体末端安全封闭以防止损伤之间,调节这一脆弱的平衡。
衰老端粒的问题
端粒起到鞋带塑料头的作用,防止染色体磨损或被误判为损伤的DNA。如果细胞将染色体末端误读为损伤,可能触发应急修复系统,导致染色体连接在一起或阻止细胞分裂。与此同时,端粒有时必须打开以便端粒酶延长它们,尤其是在分裂频繁的干细胞中。生物学家长期以来设想端粒在一种“闭合”的受保护状态与一种允许端粒酶进入但带来短暂损伤信号的“开放”状态之间切换。是否这两种状态反映了端粒的不同物理构象,或是真正独立的控制机制,一直是一个未解的问题。
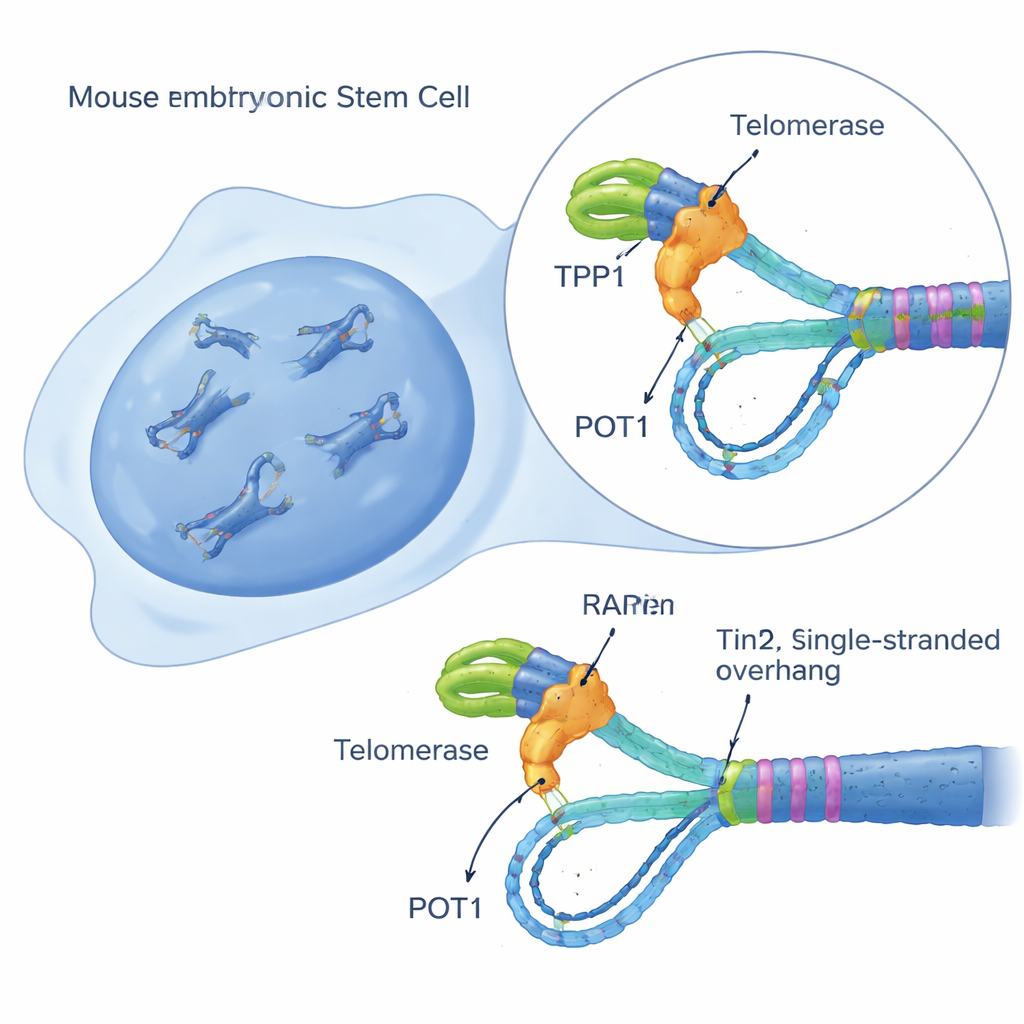
构建单细胞端粒酶活性传感器
为了解开这一谜题,研究者在小鼠胚胎干细胞中创建了一种灵敏的活细胞检测系统,称为 iTAP(可诱导端粒酶活性探测)。他们对细胞进行工程改造,仅在给予药物多西环素时产生稍作改变的端粒酶RNA模板。该“突变”模板使端粒酶在端粒上添加可与正常端粒DNA区分的修饰序列。通过荧光探针、DNA印迹和一种富集端粒片段的测序方法,研究团队能够逐细胞、逐位点地直接观察端粒酶何时何地活跃。关键是,该系统本身并不损害细胞生长或诱发损伤反应,从而允许在其他条件正常的细胞内对端粒酶行为进行干净的测量。
TPP1 招募端粒酶,POT1 则予以抑制
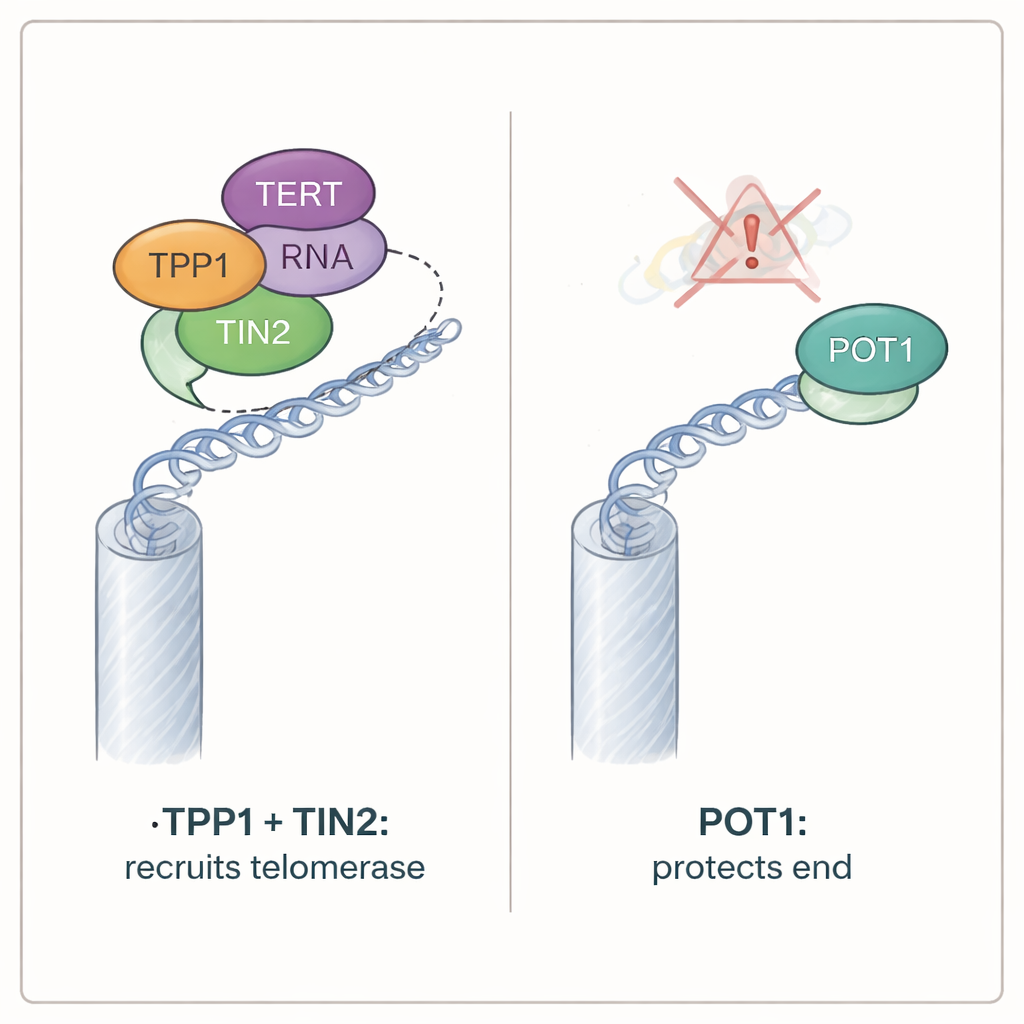
借助 iTAP,作者测试了两种关键端粒蛋白TPP1和POT1如何影响端粒酶作用。两者都属于被称为 shelterin 的更大复合体,覆盖在端粒上。当他们使用基因编辑工具去除TPP1时,突变端粒序列几乎消失,尽管突变的端粒酶RNA仍保持正常水平产生。进一步实验表明,TPP1必须与另一个shelterin成分TIN2物理连接才能招募端粒酶;破坏TPP1与TIN2的连接会关闭端粒酶活性。令人惊讶的是,破坏TPP1与POT1的相互作用并没有类似效应,而直接删除POT1本身也并未减少端粒酶的作用。事实上,缺失POT1的细胞显示出加入的突变序列量略有增加,这表明POT1通常充当刹车——限制端粒酶与端粒接触的频率,而不是帮助其招募。
POT1 守护端点,而 TPP1 在某些情况下可有可无
尽管POT1对端粒酶的招募并非必需,但它对保护染色体末端至关重要。小鼠细胞中同时缺失两种POT1变体的细胞迅速停止增殖,并表现出暴露端粒的所有典型特征:DNA损伤标志在染色体末端积聚,称为ATR的损伤感应酶被激活,染色体末端发生融合。相比之下,缺失TPP1的细胞继续生长,很少显示端粒受损的迹象,尽管它们丧失了正常的端粒酶活性。额外测试表明,POT1的防御作用依赖于其结合端粒单链突出端的能力。无法抓住该暴露DNA的POT1突变体即便还能与TPP1相互作用,也无法抑制损伤信号。相反,能够结合突出端但不再与TPP1结合的POT1变体仍能保护染色体末端。
端粒控制的新模块化视角
这些发现推翻了端粒在单一“开放”状态(允许端粒酶进入)与单一“闭合”状态(用于保护)之间切换的简单图景。相反,研究显示端粒酶招募与末端保护由可分离的分子回路控制。TPP1通过TIN2作为将端粒酶带到端粒的主要通道,而POT1则独立守护染色体末端的暴露DNA并抑制损伤反应。对非专业读者来说,结论是:细胞并不依赖染色体末端的单一开关。相反,它们使用分离但协调的旋钮——一个用于允许端粒酶进入,另一个用于保持末端安全——以维持终生的基因组稳定性。
引用: Sandhu, R., Tricola, G.M., Lee, S.Y. et al. Control of telomerase recruitment and end protection by independent shelterin components. Nat Commun 17, 1733 (2026). https://doi.org/10.1038/s41467-026-68433-0
关键词: 端粒, 端粒酶, 干细胞, DNA 损伤, 染色体保护