Clear Sky Science · zh
通过自组织机械因子活性梯度对收缩性的空间模式化是果蝇原肠胚形成的基础
组织如何自我塑形
早期胚胎能够将简单的细胞层折叠成复杂的三维结构。这项研究探讨了这一“魔术”背后的基本问题:细胞如何协调在哪里收缩和弯曲,以便组织在正确的位置和方向折叠?通过对果蝇胚胎进行高分辨率观察,作者发现了一种类似“机械形态发生素”的化学信号,它直接为细胞自身牵拉的部位建立模式,从而驱动发育过程中的关键折叠事件。
一股推动胚胎折叠的波
在果蝇 Drosophila 中,早期发育的关键步骤之一是原肠胚形成,即一层扁平的细胞内陷开始构建肠道。在胚胎的后端,一片称为后内胚层原基的细胞通过其上端(顶端)收缩率先发起该过程。最初的弯曲触发了一股穿过相邻细胞并向前推进的组织内陷波。作者表明,这一波依赖于一种名为 Fog 的信号分子——一种被分泌的因子,它激活一类称为 GPCR 的受体,进而开启由马达蛋白 Myosin-II 组成的细胞收缩机械。
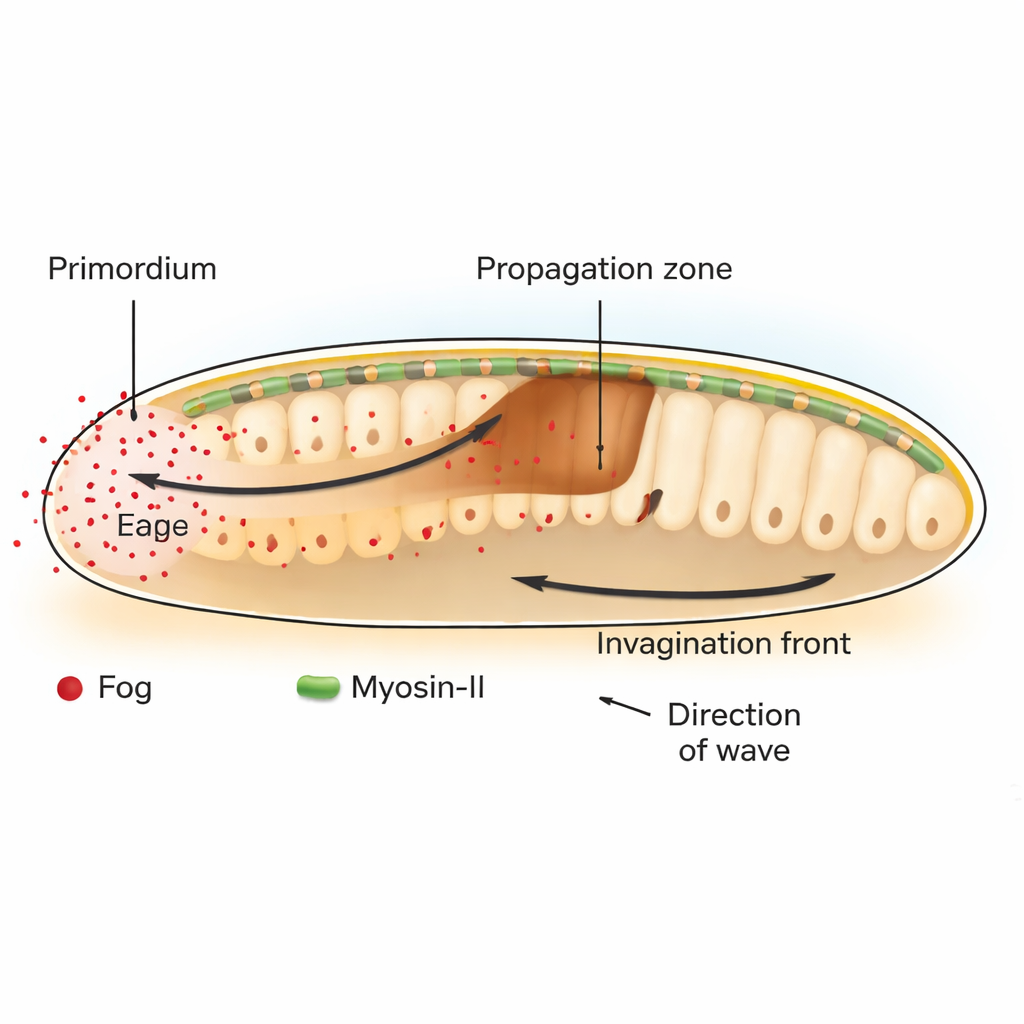
塑造力而非命运的化学物质
传统上,发育生物学家关注“形态发生素”——扩散分子,其浓度梯度指示细胞应采取的身份。在这里,Fog 的行为有所不同。它不是改变区域将成为什么类型的细胞,而是直接调节细胞收缩的强度。通过有选择地在特定区域恢复或阻断 Fog 及其下游信使 Gα,研究者证明了原基中产生的 Fog 对诱发 Myosin-II 激活并在相邻的传播区远程引发组织折叠既是必需的又是充分的。当 Fog 被固定使其不能扩散时,仅产生者细胞发生收缩,内陷波在仅一两排细胞后就停止。
细胞表面上的隐性梯度
一种自然的假设是 Fog 在细胞外形成经典的浓度梯度。利用荧光标记的 Fog 以及灵敏的荧光测量,团队却发现了相反的情况:在组织与其包膜(卵壳膜)之间的薄液层中,Fog 均匀分布。然而 Myosin-II 活性明显形成一个陡峭的传播梯度,在前进的折痕正前方最强,并在几倍细胞直径外衰减。缺失的一环是 Fog 并不是作为简单的体积浓度提示发挥作用;相反,一小部分 Fog 暂时被捕获在细胞表面,结合其 GPCR 受体,形成表面结合的活性梯度,即便游离的 Fog 分子在液体中仍然均匀混合。
受体、内吞与粘附共同塑造波
作者表明,这种表面结合的 Fog 与其受体一起自组织为一种图案。在 Fog 多次与受体接触的部位,它促进受体聚集与激活,进而招募更多的 Myosin-II。细胞的“清理”过程——GPCR 的内吞作用——随后移除活性复合体,防止失控激活并调节信号扩散的距离。当该内吞作用在遗传学上被减慢时,Myosin-II 梯度会变得更高且更宽。另一个关键角色是一组称为整合素的粘附分子。随着组织弯曲,折痕前方的细胞会压向卵壳膜;这种接触局部激活整合素,反过来增强 Fog–GPCR 信号的效力。强烈整合素参与的区域与 Myosin-II 梯度的范围相吻合,若缺失整合素,即使 Fog 信号被增强,波也会变得微弱且传播距离短。
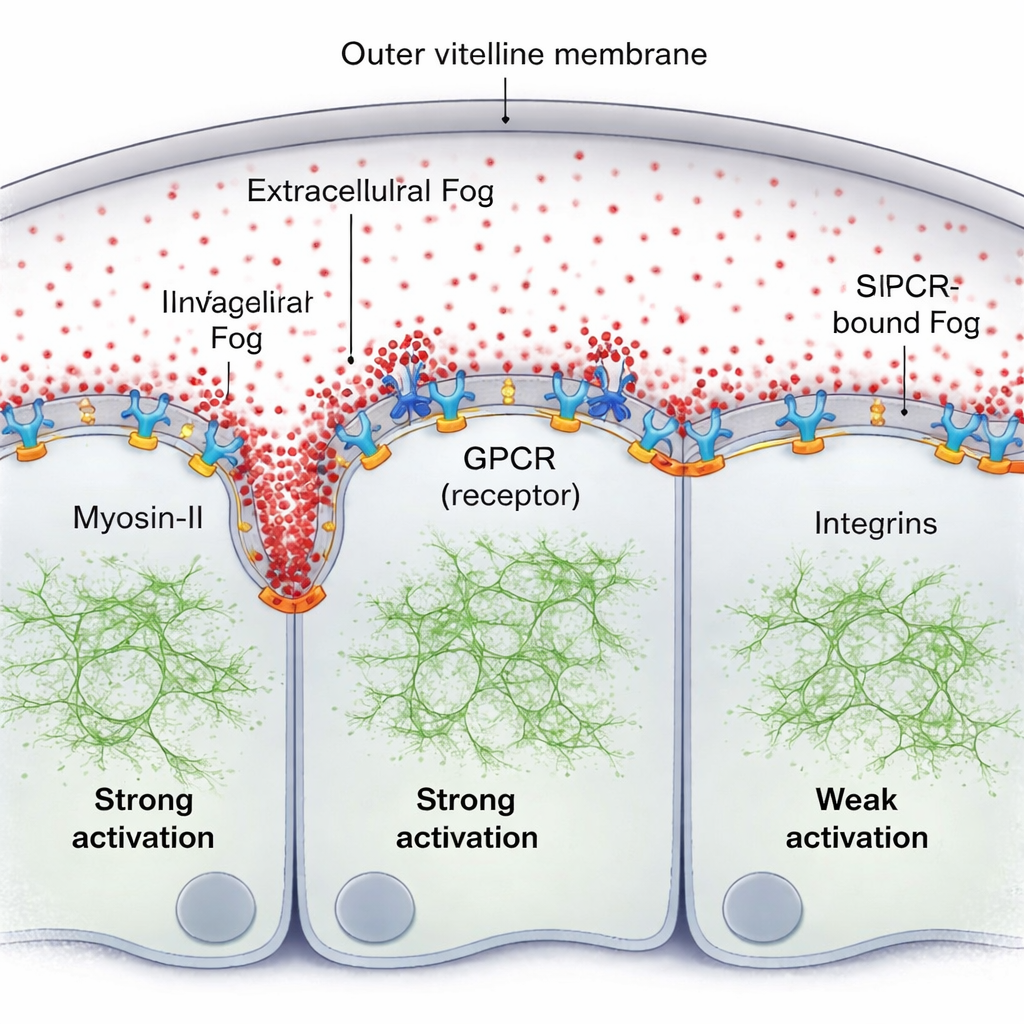
一种自我更新的机械模式
对非专业读者而言,主要结论是胚胎并不完全依赖预先绘制的化学地图来决定在哪里折叠。相反,它动态构建出一个移动的机械活性图案。Fog 虽然均匀扩散,但其有效作用在细胞表面被受体聚集、受体移除以及随着细胞与周围环境接触而变化的组织几何共同雕塑。由此产生了一种自组织的、向前传播的收缩力梯度,协同将组织向内牵拉。该工作提供了长久被理论化的“机械因子”的一个具体实例:一种扩散分子直接塑造组织中的物理力学,帮助解释生物体在发育过程中如何如此可靠地自我塑形。
引用: Mundhe, G., Dunsing-Eichenauer, V., Philippe, JM. et al. Spatial patterning of contractility by a self-organized mechanogen activity gradient underlies Drosophila gastrulation. Nat Commun 17, 1721 (2026). https://doi.org/10.1038/s41467-026-68418-z
关键词: 原肠胚形成, 机械因子, 果蝇胚胎, 组织力学, Fog 信号