Clear Sky Science · zh
哈尔丹法则通过 X:常染色体不相容性在秀丽隐杆线虫/黑线虫杂交中的作用
为什么有些杂交后代失败而有些却能存活
当密切相关的物种交配时,后代常常虚弱、不育或根本不能发育。生物学家长期观察到这些失败中一个奇特的模式:携带两种不同性染色体的性别(例如哺乳动物的 XY 雄性)通常受影响更大。这个被称为哈尔丹法则的模式,有助于解释新物种如何保持隔离。在这项研究中,研究人员使用微小的线虫揭示了这一法则的一个出人意料的简单原因:一个性染色体与其余基因组之间的不匹配。
关于杂交后代的旧难题
哈尔丹法则最早在一个多世纪前被描述,但其遗传学原因仍不明朗。两种主要观点占主导地位。一种认为性染色体进化迅速,积累了仅在杂交体中显现的隐性遗传冲突。另一种则提出雄性性状,尤其是与生育力相关的性状,变化异常迅速,以至于杂交雄性无法产生功能性精子。这两种理论都预测“异配合”性(即拥有不匹配性染色体的性别,XY、ZW 或 XO)会遇到问题,但都没有指出究竟是哪些 DNA 片段应该被指责。
打破然后重塑规则的线虫
作者转而研究两种密切相关的线虫,Caenorhabditis briggsae 和 C. nigoni,它们可以交配但通常产生虚弱或不育的雄性。当一只 C. nigoni 雌性与一只 C. briggsae 雄性交配时,具有两条 X 染色体(XX)的雌性健康,但具有单条 X(XO)的雄性不育。在相反的杂交中,XO 雄性在胚胎期就死亡。XX 健康与 XO 失败之间的这一鲜明对比使这些线虫成为研究哈尔丹法则的强有力模型。 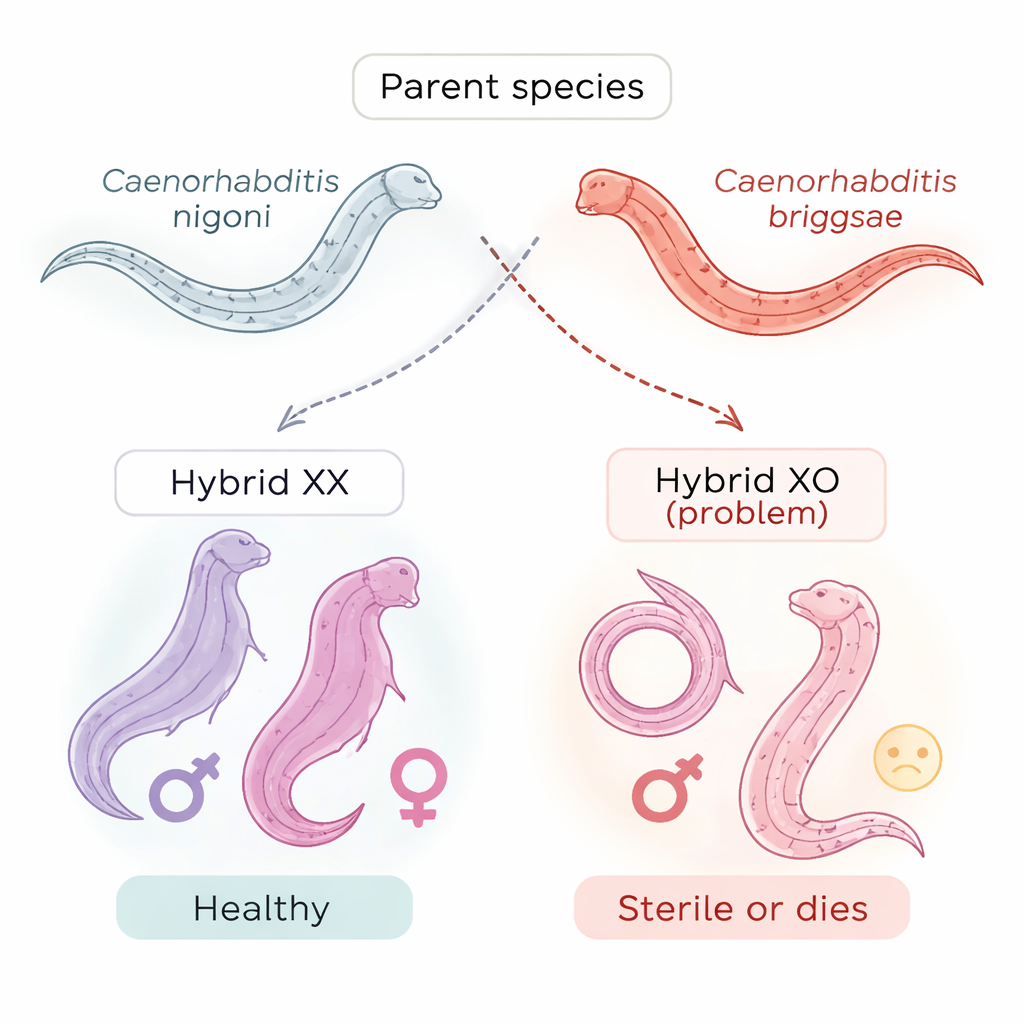
检验是否“成为雄性”真的是问题所在
为了判断是否是雄性特异基因导致了问题,研究人员利用关键性别决定基因 tra-1 的突变,强制基因上为 XX 的杂交体发育为雄性。如果雄性发育程序本身在两种物种间不兼容,这些 XX 雄性应当是不育或发育畸形的。相反,大多数发育出正常的雄性体型,产生精子,甚至能产生后代。这表明构成雄性的基本遗传程序——体型、行为和生育能力——在两种基因组结合时仍能正常工作。因此,问题并不仅仅是“雄性基因”进化得太快。
通过增加额外染色体修复杂交体
研究组接着探查真正的困难是否出在 XO 杂交体中那条单独的 X 染色体与其他染色体(即常染色体)之间的相互作用。为此他们产生了具有四套染色体而不是两套的线虫——四倍体。在这些四倍体杂交体中,雄性从每个物种继承一条 X 染色体,同时还有额外的常染色体。值得注意的是,这些四倍体杂交雄性健康,产生大量精子并能成为父本,这与不育或死亡的二倍体 XO 雄性形成鲜明对比。该结果指向二倍体杂交体中的一个具体问题:来自一个物种的孤立 X 试图与来自两种物种的混合常染色体成对工作。
基因剂量与平衡如何出错
作者提出,基因位点(位于 X 染色体还是常染色体)上的细微变化,加上用于平衡 X 上基因活性的机制(称为剂量补偿),可以在 XO 杂交体中打破基因表达的平衡。在进化过程中,每个物种微调其基因活性,使 X 连锁基因与常染色体基因协同工作。但在继承来自一个物种的单条 X 和来自两者的常染色体的杂交体中,某些基因相对于其配对伙伴会过度活跃或活性不足。这些不匹配表达水平的累积效应会扰乱异配合性别的发育,导致不育或死亡,而 XX 杂交体在很大程度上仍保持平衡。 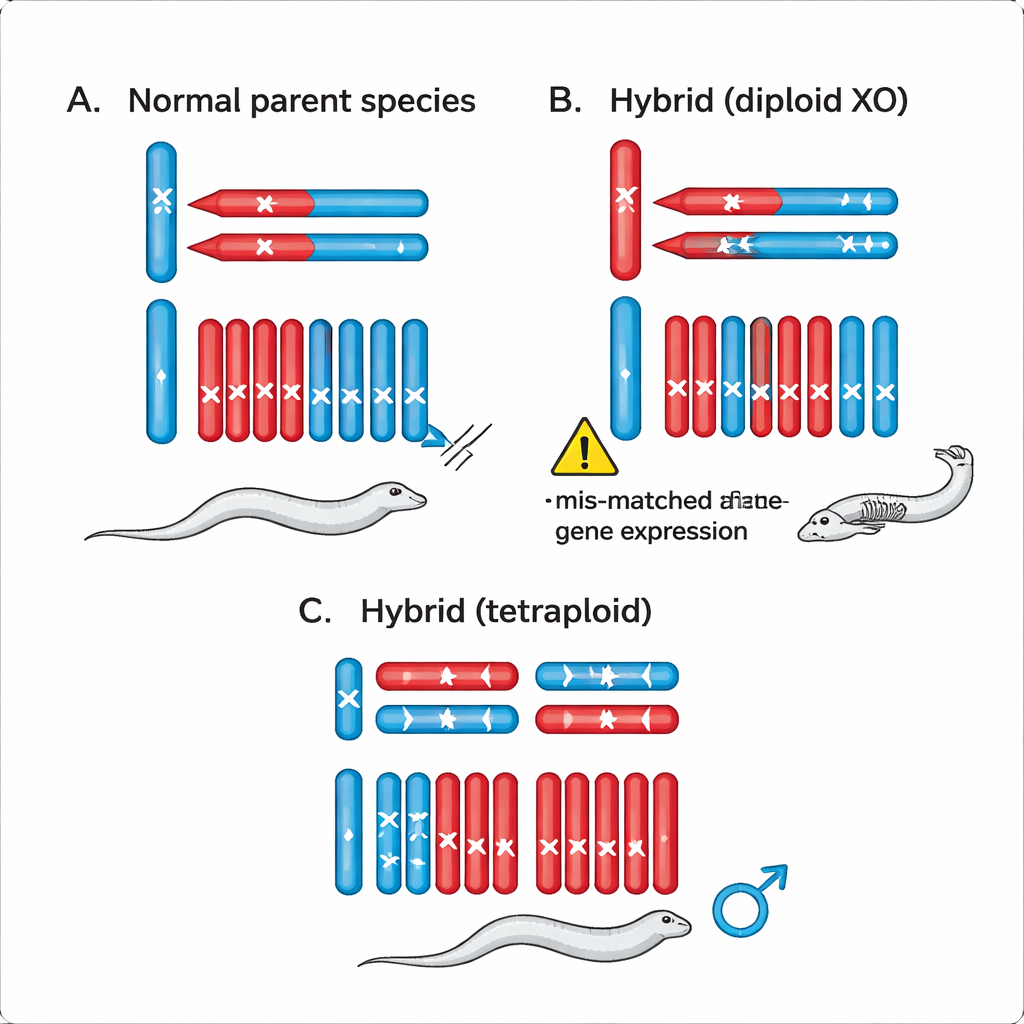
这对物种形成意味着什么
通俗地说,这项研究表明,杂交失败往往归结为一个性染色体与其余基因组之间协调不佳,而不是内在脆弱的雄性体制。在这些线虫中,一旦研究者要么重新接线性别决定机制,要么增加额外的染色体拷贝,杂交雄性就能存活并繁衍。这支持了一个简单且普遍的哈尔丹法则观点:当物种分化时,基因在 X 染色体与常染色体之间的分布和调控方式发生的细微变化会悄然累积。这些变化在各自物种内部无害,但在基因组混合时会引发问题,从而帮助巩固新兴物种之间的界限。
引用: Harbin, J.P., Shen, Y., Abubakar, A.H. et al. Haldane’s law works through X:Autosome incompatibility in Caenorhabditis briggsae/C. nigoni hybrids. Nat Commun 17, 1679 (2026). https://doi.org/10.1038/s41467-026-68383-7
关键词: 杂交不育, 性染色体, 哈尔丹法则, 物种形成, 线虫遗传学